Хлоропласты зеленые пластиды. Какие пластиды имеют пигмент хлорофилл. Отдел Сине-зеленые водоросли. Особенности организации, жизненные формы, пигменты, жизненный цикл
ПЛАСТИДЫ
Пластиды всегда находятся в протоплазме, близки к ней по физическим и химическим свойствам, возникают только от пластид. Они способны к росту и размножаются делением, могут образовывать в своем теле (в строме) определенные пигменты и формировать внутри стромы крахмал. В зависимости от содержания тех или иных пигментов находится окраска и функции основных пластид высших растений: а) зеленых пластид (хлоропластов), б) красных и желтых (хромопластов) и в) бесцветных (лейкопластов).
Все пластиды, по-видимому, имеют сходное строение; лучше изучено строение хлоропластов.
1 - клетка из нити Zygnema cruciatum ; 2 - отдельная клетка из нити спирогиры; п - пиреноиды.
х - хроматофор; п - пиреноиды с крахмалом; я - ядро.
Хлоропласты . Хлоропласты высших растений (называемые также хлорофилловыми зернами) по форме до некоторой степени сходны с линзами: в плане хлоропласт имеет очертание, более или менее близкое к кругу, а при рассматривании в профиль напоминает эллипс. Если хлорофилловые зерна лежат тесным слоем, то, нажимая друг на друга, они принимают угловатую форму. Число хлорофилловых зерен в различных клетках очень изменчиво. Например, в клетках листа клещевины количество хлорофилловых зерен колеблется от 10 до 36, в клетках Elodea densa - от 26 до 32. Диаметр хлорофилловых зерен составляет 4-9 μ.
По своим размерам хлорофилловые зерна менее разнообразны, чем ядра, а тем более сами клетки, хотя некоторые авторы отмечают, что хлорофилловые зерна крупнее в клетках больших размеров. Когда И. И. Герасимов в культурах спирогиры получал крупные клетки с двойной массой ядра, то в таких клетках и спиральные хлоропласты были крупнее, чем в нормальных, и число их возрастало с 8 до 12-13 (рис. 18). Исключительно крупные хлоропласты отмечены для Peperomia metallica : диаметр
хлоропластов достигает 24 μ, но здесь следует отметить, что число их в клетке очень невелико - их только 4.
Хлоропласты могут изменять форму и размеры. Некоторые изменения зависят от поверхностного натяжения; при возрастании его уменьшается величина поверхности, и форма пластиды приближается к сферической - пластида "округляется"; при уменьшении поверхностного натяжения пластида удлиняется. Форма хлоропластов может меняться в зависимости от освещения; например, в листьях клещевины, подвергнутых затенению, хлоропласты становятся почти изодиаметрическими (с наибольшим размером ∼6,3 μ и наименьшим ∼5,7 μ); на ярком свету они меняют форму на чечевицеобразную (с диаметром ∼8,3 μ и толщиной ∼3,6 μ).
В культуре водяной чумы (элодеи) при разных температурах хлорофилловые зерна в листьях, выросших при более высокой температуре, получались почти вдвое меньшими.
Очень разнообразны по форме хлоропласты водорослей - хроматофоры (рис. 28, 29).
В окрашенных пластидах - хроматофорах - многих водорослей (рис. 28, 29) и некоторых из печеночников, относящихся к роду Anthoceros , имеются особые, тягуче-жидкие тельца, чаще всего округлой или угловатой формы; эти тельца, называемые пиреноидами , богаты белковыми веществами, но нуклеинов не содержат. Вокруг пиреноидов обычно располагаются мелкие крахмальные зерна; эти зерна крахмала образуются в клетке в первую очередь, а исчезают в последнюю. Пиреноиды образуются путем деления уже существующих, но могут и возникать в клетке заново.
Хлоропласты содержат в строме четыре пигмента: два зеленых (хлорофилл a и хлорофилл b ), оранжево-красный (каротин, или, иначе, каротен) и желтый (ксантофилл).
По своему химическому составу хлорофилл представляет собой сложный эфир дикарбоновой кислоты хлорофиллина и двух спиртов - метилового и фитола.
Хлорофилл а отличается от хлорофилла b по количеству атомов водорода и кислорода.
Хлорофилл а имеет синеватый оттенок, хлорофилл b - желтоватый. Молекулярный вес хлорофилла равен ∼ 900.
И. П. Бородин, обрабатывая срезы зеленых частей растения на предметном стекле этиловым спиртом, получал после медленного высушивания препарата темно-зеленые или почти черные кристаллы в виде трех- или шестиугольных пластинок и тетраэдров. В дальнейшем было выяснено, что это кристаллы хлорофилла, в молекулах которого фитольная группа замещена этильной.
Центральное место в молекулах хлорофиллов a и b занимает атом магния, связанный с 4 атомами азота.
В одном хлорофилловом зерне содержится 6% хлорофилла; остальное составляют вода, белки, липиды и др.
Из пигментов, сопровождающих хлорофиллы, оранжево-красный каротин представляет ненасыщенный углеводород формулы C 40 H 56 , а желтый ксантофилл (C 40 H 56 O 2) - двухатомный спирт, как бы продукт окисления каротина. Каротин и ксантофилл относятся к обширной группе каротиноидов - пигментов желтого, оранжевого и красного цветов, ряд других представителей которых также встречается в растениях. Всю совокупность пигментов хлоропласта иногда называют "хлорофиллом" в широком смысле слова.
Вытяжка зеленых листьев и в меньшей мере сами листья обнаруживают явление флуоресценции. В проходящем свете вытяжка кажется зеленой, а в падающем - вишнево-красной. За очень редкими исключениями, в органах покрытосеменных растений при развитии их в отсутствии света зеленые пигменты в хлоропластах не образуются или образуются в ничтожном количестве; на свету происходит их быстрое позеленение.
В хлоропластах совершается сложный процесс фотосинтеза - образования углеводов из углекислого газа и воды под действием энергии солнечного света .
Окончательный результат процесса фотосинтеза можно представить в виде следующей реакции:
6CO 2 + 6H 2 O + 674 ккал → C 6 H 12 O 6 + 6O 2
Образовавшийся углевод обычно полимеризуется в крахмал по схеме:
n C 6 H 12 O 6 → (C 6 H 10 O 5) n + n H 2 O

Рис. 30. Хлорофилловые зерна листа мха Funaria hydrometrica
:
1 - участок взрослого листа с несколькими клетками (в плане); в постенном слое протоплазмы расположены хлорофилловые зерна с мелкими крахмальными зернами (отмечены белым цветом); 2-10 - отдельные хлорофилловые зерна с крахмалом: 2 - молодое, 3 - более взрослое, 9 и 10 - делящиеся, 4, 5 и 6 - заполненные крахмалом, 7 - молодое, набухшее в воде, 8 - расплывшееся в воде и оставившее после себя крахмальные зерна.
Крахмал откладывается в хлоропластах в виде мелких зерен ассимиляционного , или автохтонного , крахмала (рис. 30, 32).
У некоторых растений, преимущественно однодольных, ассимиляционного крахмала обычно не образуется (кроме как в замыкающих клетках устьиц) и продуктом фотосинтеза является глюкоза. При сильно повышенном содержании CO 2 в атмосфере в хлоропластах сахарообразующих растений (сахарного тростника, сахарной свеклы) на свету появляется крахмал.
Способность к фотосинтезу и ряд других свойств пластид как биологически активных систем объясняются наличием в хлоропластах ферментов . Эта очень сложная ферментативная система обеспечивает не только

весь процесс фотосинтеза, но и отток продуктов ассимиляция из хлоропласта. В состав этой системы входит зеленый пигмент пластид - хлорофилл.
О внутренней структуре хлоропласта было очень много споров, выдвигалось много теорий, но только применение электронного микроскопа дало возможность более детально изучить его субмикроскопическое строение (рис. 31, 32). В настоящее время считают, что хлоропласты высших растений имеют пластинчатую структуру. Пластинки стромы чередуются с пластинками, состоящими из гранул (зернышек), содержащих хлорофилл. Связь между гранулами и стромой в настоящее время еще не совсем ясна.
В выяснении вопросов роли хлорофилла в процессе фотосинтеза и значении лучей различных областей солнечного спектра в этом процессе большая заслуга принадлежит К. А. Тимирязеву. Тимирязев изучал хлорофилл как "связующее звено между солнцем и жизнью", а хлорофилловое зерно - как тот фокус, ту точку в мировом пространстве, где солнечный луч, превращаясь в химическую энергию, становится источником всей жизни на Земле.
Тимирязев установил, что наиболее интенсивно поглощаются красные лучи (с длиной волны от

Справа - сформированный хлоропласт, в строме которого видны три линзовидных крахмальных зерна.
730 до 680 м μ) и в несколько меньшей мере лучи сине-фиолетовой части спектра (с длиной волны 470 м μ и меньше).
Осенью перед опадением листа хлорофилловые зерна желтеют; это пожелтение зависит от того, что зеленые пигменты разрушаются раньше, чем желтые; пожелтение связано с оттоком веществ из листовой пластинки в осевые органы.
Хромопласты . Хромопласты - пластиды, содержащие пигменты из числа каротиноидов (каротин и ксантофилл). Они имеют окраску от желтой (в лепестках лютиков) и оранжевой (в кожуре апельсинов) до оранжево-красной (в корнях моркови) и ярко-красной (в плодах шиповников).

Рис. 33. Клетка чашелистика настурции Tropaeolum majus
с ядром и хромопластами.

1 - в клетках кожицы Philodendron grandifolium; 2, 3 - в клетках семени Melandrium macrocarpum; 4 - в клетках корня Phajus grandifolius (изображено лишь ядро с лейкопластами подле него).
По форме хромопласты весьма разнообразны. Изредка они бывают эллипсоидальными или при тесном расположении многоугольно-таблитчатыми, лопастными и т. д. Обычно хромопласт имеет игловидное и угловатое очертание, его строма растянута пигментом, составляющим преобладающую по объему часть хромопласта (рис. 33). У многих растений в хромопластах отмечается наличие крахмала.
В хромопластах иногда обособляются белковые вещества или образуются капли масла: в клетках кожуры плодов апельсина и других цитрусовых пигмент хромопластов частично растворен в эфирных маслах.
Не всегда окраска плодов зависит только от хромопластов. От наличия пластид зависят цвета зеленый, желтый, кирпично-красный (лепестки цветков лютиковых, сложноцветных, корни моркови, многие созревающие плоды). Цвета же синий, малиновый, темно-красный (плоды малины, калины) зависят от окраски клеточного сока, содержащего антоцианы. Часто получается смешанный цвет, обусловленный окраской клеточного сока и пластид.
Очень показательно провести отделение каротина бензолом от спиртовых вытяжек плодов красного перца и рябины. Если к спиртовой вытяжке плодов перца добавить бензол, то сверху в бензоле окажется растворенным каротин, а ксантофилл окажется внизу в спирте. Клеточный сок (также остается в спирте) бесцветный. Если повторить этот опыт с плодами рябины, можно видеть, что каротина в них не так уж много, так как бензольная фракция будет гораздо более бледной, чем таковая красного перца, а спирт останется окрашенным в розовый цвет от наличия антоциана.
Белый цвет лепестков обусловлен равномерным рассеиванием света при отражении его от пузырьков воздуха, заключенных в сильно развитых межклеточных пространствах и на поверхности органа.
Желтый цвет некоторых цветков и плодов (георгин, льнянка, мак, лимон) зависит также от пигмента клеточного сока, родственного антоциану, - антохлора.
Окраска частей растений может зависеть еще от цвета отмерших клеток и их измененного содержимого (оболочка семян, поверхность стволов деревьев).
Лейкопласты . Пластиды, не содержащие в строме пигментов и называемые лейкопластами , имеются во многих клетках большинства растений (рис. 34). Так как лейкопласты бесцветны и к тому же преломляют свет почти так же, как протоплазма, не всегда легко обнаружить их присутствие в клетке.
По форме лейкопласты обычно почти шаровидны. В тех случаях, когда в их строме находится крахмал или белок, они принимают иные очертания. Сравнительно богаты лейкопластами образовательные ткани, подземные органы, семена.
В лейкопластах может образовываться крахмал, отлагающийся в виде зерен в их строме.
Во многих случаях крахмал накопляется в лейкопласте в столь большом количестве, что живое тело пластиды (строма) оттесняется на периферию. Его можно не без труда заметить в виде очень тонкой пленки на поверхности крахмального зерна; в этих случаях лейкопласт является крахмалонакопителем в полной мере (подробнее о запасном крахмале см. на стр. 78).
Движение пластид . Кроме перемещений пластид, связанных с распределением их между дочерними клетками, образующимися в результате деления клетки, совершаются другие передвижения их: 1) зависящие от возрастных изменений клетки и пластид и 2) обратимые и многократно повторяющиеся движения, связанные с изменениями в направлении и интенсивности воздействия факторов среды ("ориентировочные" движения пластид).
Хлорофилловые зерна, находящиеся в постенном слое протоплазмы, могут перемещаться в клетке. Они не только пассивно увлекаются протоплазмой при ее движении, но, в зависимости от силы и направления света, сами могут менять свое положение в клетке. И. П. Бородин показал, что лучше всего это видно на тонких листочках ряски (Lemna trisulca ).
На рисунке 35 изображено размещение хлорофилловых зерен в клетках, которые находились в различных условиях освещения. На рассеянном свету хлорофилловые зерна располагаются по стенкам, которые находятся под прямым углом к главному направлению лучей; на каждое зерно непосредственно попадает бо́льшая часть падающего света. На ярком солнечном свету хлорофилловые зерна перемещаются на боковые стенки, которые лежат
вдоль падающих лучей. Свет, проникающий в клетку, не попадает на зерно прямо, а рассеивается в стороны и освещает при этом зерна более равномерно. В темноте зерна располагаются по стенкам, которые соприкасаются со стенками соседних клеток, как говорят по теплым стенкам, но последнее расположение не всегда удается наблюдать.
Интересно, что освещение отдельных хлорофилловых зерен происходит весьма равномерно. Самостоятельная скорость перемещения хлорофилловых зерен в клетке равна 0,12 μ в секунду, т. е. скорость значительно меньшая, чем движение амебы и плазмодия.
На расположение хлорофилловых зерен влияет не только сила и направление света. В некоторых случаях наблюдалось перемещение хлорофилловых зерен к тем участкам клеточной стенки, к которым подходят межклеточные пространства; по межклетникам поступает углекислота, поглощаемая в процессе ассимиляции.

Рис. 35. Перемещение хлорофилловых зерен в клетках ряски трехдольной (Lemna trisulca
):
А - часть растения в плане, верхняя левая часть находилась в темноте, правая нижняя часть - на рассеянном свету (по Бородину); Б - частичные поперечные разрезы через растения, выдержанные: 1 - на рассеянном свету; 2 - на прямом солнечном; 3 - в темноте.
Происхождение пластид . Резких границ между пластидами различных категорий нет. Пластиды одного типа могут переходить в пластиды другого типа. В самом деле, в проростках и во взрослых особях многие из пластид, возникающих в результате деления лейкопластов клеток зародыша, становятся хлоро- и хромопластами. Хлоропласт, теряя хлорофилл и обогащаясь каротиноидами (как в созревающих плодах шиповника), становится хромопластом; лейкопласт при накоплении в его строме каротиноидов превращается в хромопласт (как в корнях моркови). Достаточно обнажить верхнюю часть растущего в почве красного "корнеплода" моркови от земли, чтобы через некоторое время эта часть гипокотиля и корня позеленела: хромопласты клеток становятся хлоропластами.
Вопрос о первоначальном возникновении пластид в клетках растений в настоящее время еще не совсем ясен. Существовала теория возникновения пластид из хондриосом; большинство исследователей от нее отказались. В последнее время высказывается мнение, что пластиды возникают из особых образований - пропластид.
В настоящее время известно, что в растениях хлорофилл присутствует в нескольких формах, но известно строение только двух его форм - a и b , причем и здесь положение двойных связей и характер связи магния с азотом еще нуждаются в уточнении.
А. С. Фаминцын впервые установил, что процесс фотосинтеза может происходить и при искусственном освещении.
Группируются в граны, которые представляют собой стопки сплюснутых и тесно прижатых друг к другу тилакоидов, имеющих форму дисков. Соединяются граны с помощью ламелл. Пространство между оболочкой хлоропласта и тилакоидами называется стромой . В строме содержатся хлоропластные молекулы РНК , пластидная ДНК, рибосомы , крахмальные зёрна, а также ферменты цикла Кальвина .
Происхождение
В настоящее время общепризнано происхождение хлоропластов путем симбиогенеза. Предполагают, что хлоропласты возникли из цианобактерий , так как являются двухмембранным органоидом, имеют собственную замкнутую кольцевую ДНК и РНК, полноценный аппарат синтеза белка (причем рибосомы прокариотического типа - 70S), размножаются бинарным делением , а мембраны тилакоидов похожи на мембраны прокариот (наличием кислых липидов) и напоминают соответствующие органеллы у цианобактерий. У глаукофитовых водорослей вместо типичных хлоропластов в клетках содержатся цианеллы - цианобактерии, потерявшие в результате эндосимбиоза способность к самостоятельному существованию, но отчасти сохранившие цианобактериальную клеточную стенку .
Давность этого события оценивают в 1 - 1,5 млрд лет .
Часть групп организмов получала хлоропласты в результате эндосимбиоза не с прокариотными клетками, а с другими эукариотами, уже имеющими хлоропласты . Этим объясняется наличие в оболочке хлоропластов некоторых организмов более чем двух мембран . Самая внутренняя из этих мембран трактуется как потерявшая клеточную стенку оболочка цианобактерии, внешняя - как стенка симбионтофорной вакуоли хозяина. Промежуточные мембраны - принадлежат вошедшему в симбиоз редуцированному эукариотному организму. У некоторых групп в перипластидном пространстве между второй и третьей мембраной располагается нуклеоморф, сильно редуцированное эукариотное ядро .
Модель хлоропласта
Строение
У различных групп организмов хлоропласты значительно различаются по размерам,строению и количеству в клетке. Особенности строения хлоропластов имеют большое таксономическое значение .
Оболочка хлоропластов
У различных групп организмов оболочка хлоропластов отличается по строению.
У глаукоцистофитовых, красных, зеленых водорослей и у высших растений оболочка состоит из двух мембран. У других эукариотных водорослей хлоропласт дополнительно окружен одной или двумя мембранами. У водорослей, обладающих четырехмембранными хлоропластами, наружная мембрана обычно переходит в наружную мембрану ядра.
Перипластидное пространство
Ламеллы и тилакоиды
Ламеллы соединяют полости тилакоидов
Пиреноиды
Пиреноиды - центры синтеза полисахаридов в хлоропластах . Строение пиреноидов разноообразно, и не всегда они морфологически выражены. Могут быть внутрипластидными и стебельчатыми, выступающими в цитоплазму. У зеленых водорослей и растений пиреноиды располагаются внутри хлоропласта, что связано с внутрипластидным запасанием крахмала.
Стигма
Стигмы или глазки встречается в хлоропластах подвижных клеток водорослей. Располагаются вблизи основания жгутика. Стигмы содержат каротиноиды и способны работать как фоторецепторы.
См. также
Примечания
Комментарии
Примечания
Литература
- Белякова Г. А. Водоросли и грибы // Ботаника: в 4 т. / Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. - М .: Издательский центр «Академия», 2006. - Т. 1. - 320 с. - 3000 экз. - ISBN 5-7695-2731-5
- Карпов С.А. Строение клетки протистов. - СПб. : ТЕССА, 2001. - 384 с. - 1000 экз. - ISBN 5-94086-010-9
- Lee, R. E. Phycology, 4th edition. - Cambridge: Cambridge University Press, 2008. - 547 с. - ISBN 9780521682770
| Органоиды эукариотической клетки | ||
|---|---|---|
 |
||
Wikimedia Foundation . 2010 .
Смотреть что такое "Хлоропласты" в других словарях:
- (от греч. chloros зелёный и plastos вылепленный), внутриклеточные органоиды (пластиды) растений, в к рых осуществляется фотосинтез; благодаря хлорофиллу окрашены в зелёный цвет. Встречаются в клетках разл. тканей надземных органов растений,… … Биологический энциклопедический словарь
- (от греч. chloros зеленый и plastos вылепленный образованный), внутриклеточные органоиды растительной клетки, в которых осуществляется фотосинтез; окрашены в зеленый цвет (в них присутствует хлорофилл). Собственный генетический аппарат и… … Большой Энциклопедический словарь
Тела, заключающиеся в клетках растений, окрашенные взеленый цвет и содержащие хлорофилл. У высших растении Х. имеют весьмаопределенную форму и называются хлорофилльными зернами; у водорослейформа их разнообразна и они называются хроматофорами или … Энциклопедия Брокгауза и Ефрона
Хлоропласты - (от греческого chloros зелёный и plastos вылепленный, образованный), внутриклеточные структуры растительной клетки, в которых осуществляется фотосинтез. Содержат пигмент хлорофилл, окрашивающий их в зелёный цвет. В клетке высших растений от 10 до … Иллюстрированный энциклопедический словарь
- (гр. chloros зеленый + lastes образующий) зеленые пластиды растительной клетки, содержащие хлорофилл, каротин, ксантофилл и участвующие в процессе фотосинтеза ср. хромопласты). Новый словарь иностранных слов. by EdwART, 2009. хлоропласты [гр.… … Словарь иностранных слов русского языка
- (от греч. chlorós зелёный и plastós вылепленный, образованный) внутриклеточные органеллы растительной клетки Пластиды, в которых осуществляется фотосинтез. Окрашены в зелёный цвет благодаря присутствию в них основного пигмента фотосинтеза … Большая советская энциклопедия
Ов; мн. (ед. хлоропласт, а; м.). [от греч. chlōros бледно зелёный и plastos вылепленный] Ботан. Тельца в протоплазме растительных клеток, содержащие хлорофилл и участвующие в процессе фотосинтеза. Концентрация хлорофилла в хлоропластах. * * *… … Энциклопедический словарь
Тела, заключающиеся в клетках растений, окрашенные в зеленый цвет и содержащие хлорофилл. У высших растений X. имеют весьма определенную форму и называются хлорофилльными зернами (см.); у водорослей форма их разнообразна и они называются… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Мн. Зеленые пластиды растительной клетки, содержащие хлорофилл, каротин и участвующие в процессе фотосинтеза. Толковый словарь Ефремовой. Т. Ф. Ефремова. 2000 … Современный толковый словарь русского языка Ефремовой
- (от греч. chloros зелёный и plastоs вылепленный, образованный), внутриклеточные органоиды растит. клетки, в к рых осуществляется фотосинтез; окрашены в зелёный цвет (в них присутствует хлорофилл). Собств. генетич. аппарат и белоксинтезирующая… … Естествознание. Энциклопедический словарь
Пластиды являются органоидами протопласта, характерными только для растительных клеток. Их нет лишь у бактерий, синезеленых водорослей и, возможно, грибов.
У высших растений пластиды находятся во взрослых вегетативных клетках всех органов - в стебле, листе, корне и цветке. Пластиды - это сравнительно крупные органоиды, значительно крупнее митохондрий, а иногда даже и крупнее ядра, более плотные, чем окружающая их цитоплазма, хорошо видимые в световой микроскоп. Они имеют характерное строение и выполняют различные функции, связанные главным образом с синтезом органических веществ.
Во взрослой растительной клетке в зависимости от окраски, формы и функции различают три основных типа пластид: хлоропласта (пластиды зеленого цвета), хромопласты (пластиды желтого и оранжевого цвета) и лейкопласты (бесцветные пластиды). Последние по своему размеру меньше пластид двух предыдущих типов.
Хлоропласты
Структурной основой хлоропласта являются белки (около 50% сухого веса), они содержат также 5-10% хлорофилла и 1-2% каротиноидов. Как и в митохондриях, в хлоропластах обнаружено небольшое количество РНК (0,5-3,5%) и еще меньше ДНК. Исключительное значение хлоропластов в том, что в них происходит процесс фотосинтеза. Крахмал, образующийся при фотосинтезе, называется первичным, или ассимиляционным, он откладывается в хлоропластах в виде мелких крахмальных зерен. Для нормального протекания фотосинтеза необходимо присутствие хлорофилла. Хлорофилл - главное действующее начало в осуществлении фотосинтеза. Он поглощает энергию света и направляет ее на совершение фотосинтетических реакций. Из пластид хлорофилл можно извлечь с помощью спирта, ацетона или других органических растворителей. Роль желтых пигментов в фотосинтезе еще недостаточно выяснена. Предполагают, что они также поглощают солнечную энергию и передают ее хлорофиллу или же вместе с ним осуществляют специфические, важные для фотосинтеза реакции.
В соответствии с их функциями хлоропласты находятся преимущественно в фотосинтезирующих органах и тканях, обращенных к свету - в листьях и молодых стеблях, незрелых плодах. Иногда хлоропласты встречаются даже в корнях, например, в придаточных корнях кукурузы. Но основное их количество сосредоточено в клетках мезофилла (мякоти) листа.
В отличие от других органоидов, хлоропласты высших растений характеризуются однообразием и постоянством формы и размеров. Чаще всего они обладают дискообразной или линзовидной формой и когда лежат плашмя, имеют округлые или многоугольные очертания. В этом случае их часто называют также хлорофилловыми зернами . Размер хлоропластов довольно постоянен и даже у разных видов высших растений колеблется в незначительных пределах, составляя в среднем 3-7 мк (толщина 1-3 мк). Более крупные хлоропласты у высших растений встречаются редко. Например, у селагинелл (плауновидных) в клетках кожицы листьев встречаются один-два крупных хлоропласта пластинчатой формы. Величина и форма хлоропластов изменяются в зависимости от внешних условий. У растений тенелюбивых хлоропласты в общем крупнее, чем у светолюбивых, и, как правило, более богаты хлорофиллом. Обычно клетка несет большое количество хлоропластов, и число их сильно меняется; в среднем же в ней насчитывается от 20 до 50 хлоропластов. Особенно богаты хлоропластами листья, а также молодые незрелые плоды. Общее количество хлоропластов в растении может быть громадным; например, во взрослом дереве насчитываются десятки и сотни миллиардов хлоропластов. Число хлоропластов в клетке связано с их величиной. Так, у кукурузы в клетках листьев обычно содержится по нескольку хлоропластов, но у сортов с особенно крупными хлоропластами число их в клетке снижается до двух.
У многих низших растений (водорослей) форма, число и размеры хлоропластов весьма разнообразны. Они могут иметь пластинчатую форму (Mougeotia), звездчатую (Zygnema) или быть в виде спиральных лент (Spirogyra) и ребристых цилиндров (Closterium). Такие хлоропласты обычно очень крупны, встречаются в клетке в небольшом количестве (от одного до нескольких) и называются хроматофорами . Но и у водорослей могут встречаться хлоропласты обычной линзовидной формы, и в этом случае число их в клетке обычно велико.
В клетках высших растений хлоропласты расположены в цитоплазме таким образом, что одна из их плоских сторон обращена к оболочке клетки, причем особенно много их около межклетников, заполненных воздухом. Здесь они тесно прилегают друг к другу и очертания их становятся угловатыми. Однако положение хлоропластов в клетке может меняться в зависимости от внешних условий и прежде всего освещенности. Они располагаются в клетке так, что улавливают свет наилучшим образом, не подвергаясь вместе с тем действию прямых солнечных лучей. В листьях некоторых растений на рассеянном свету хлоропласты располагаются преимущественно на тех стенках клеточной оболочки, которые обращены к поверхности органа, на ярком же свету они сосредоточиваются на боковых стенках или поворачиваются к лучам узкой стороной, т. е. ребром. Такое же передвижение хлоропластов наблюдается иногда и под влиянием других раздражителей - температурных, химических, механических и др. Является ли перемещение пластид активным или пассивным (током цитоплазмы), до конца еще не выяснено, но в настоящее время больше аргументов в пользу активного перемещения.
Исходя из сложности процессов фотосинтеза, состоящего из целого ряда реакций, каждая из которых катализируется особым ферментом, можно предположить, что хлоропласты имеют упорядоченную и сложную структуру. И действительно, уже в обычном световом микроскопе часто видно, что хлоропласты не являются совершенно гомогенными, а в них наблюдаются более темные мелкие зернышки, ориентированные параллельно поверхности хлоропласта, которые были названы гранами . Исследования с помощью электронного микроскопа подтвердили существование гран и показали, что весь хлоропласт в целом и граны имеют сложную структуру.
Как и митохондрии, хлоропласты представляют собой мембранные структуры, свободно лежащие в цитоплазме. От цитоплазмы они отграничены двухмембранной оболочкой с ясно видимым светлым промежутком между мембранами. Мембраны эти, как предполагают, являются гладкими и не содержат прикрепленных частиц. До недавнего времени считали, что оболочка хлоропластов сплошная, не имеет отверстий и что ее мембраны не соединены с мембранами эндоплазматической сети. Но сейчас накапливаются данные, показывающие, что это не всегда так. Иногда в отдельных местах оболочки могут возникать субмикроскопические отверстия. Возможно, что в отдельные периоды деятельности хлоропластов эти «поры» состоят в тесном контакте с эндоплазматической сетью, однако этот контакт кратковременен. Оболочка хлоропласта, обладая свойством избирательной проницаемости, играет регулирующую роль в обмене веществ между цитоплазмой и хлоропластом.
Тело хлоропласта пронизано системой двухмембранных пластин, называемых ламеллами . Пространство между ламеллами заполнено водянистой белковой жидкостью - стромой , или матриксом хлоропласта. В строме могут находиться крахмальные зерна, капли масла и рибосомоподобные частицы. Совсем недавно с помощью особо тонких методов приготовления препаратов в строме хлоропластов некоторых растений были обнаружены скопления параллельных фибрилл диаметром 80-100 Å и длиной свыше 1000 Å. Эти пучки микрофибрилл хлоропластов получили название стромацентров . Их функция совершенно не выяснена.
В отдельных участках хлоропласта ламеллы довольно плотно подходят друг к другу, располагаясь параллельно его поверхности, в результате чего в этих участках образуются скопления ламелл, называемые гранами. Внутри граны парные мембраны ламелл сливаются по краям, образуя замкнутые уплощенные мешочки, называемые дисками , или тилакоидами . Пачки таких дисков и образуют грану. Отдельные граны связаны между собой в единую систему с помощью ламелл, пронизывающих межгранные пространства. Хлорофилл не рассеян диффузно по хлоропласту, а сосредоточен в ламеллах, как предполагают, в виде мономолекулярного слоя. Рибосомы находятся не только в матриксе, но могут встречаться и на поверхности гран.

Число дисков в гранах колеблется от двух до нескольких десятков, а диаметр в зависимости от вида растения - от 0,3 до 2 мк. Поэтому у многих растений граны не видны в световой микроскоп. Число и расположение гран в хлоропласте зависят от вида растения, возраста и активности хлоропластов. В хлоропластах Aspidistra гран так много, что они соприкасаются между собой, а у так называемых клеток-спутников тыквы основной объем хлоропласта занят стромой. В хлоропластах листьев томата и хризантемы граны беспорядочно разбросаны, а в хлоропластах табака они правильно ориентированы по отношению к поверхности хлоропласта и располагаются на равном расстоянии друг от друга. У светолюбивых растений граны мельче, чем у теневыносливых.
Структура хлоропластов высших растений прекрасно приспособлена к выполнению их главной функции - фотосинтеза. Уже само разделение хлорофиллоносного аппарата на мелкие пластиды означает громадное увеличение активной поверхности. За счет образования мембран и гран эта поверхность увеличивается еще более. Большая активная поверхность и тонкая пространственная ориентация обеспечивают легкий доступ энергии кванта света и возможность переноса этой энергии к химическим системам, участвующим в фотосинтезе. Принцип замкнутых камер - тилакоидов, благодаря пространственному разобщению позволяет одновременно и независимо осуществлять один и тот же комплекс реакций, составляющих фотосинтез. В рибосомах хлоропластов идет синтез белка.
В клетках некоторых водорослей (спирогира) и реже высших растений (например, клетки так называемой обкладки проводящих пучков у кукурузы) встречаются безгранные хлоропласты, у которых ламеллы пронизывают строму, не образуя отчетливых гран.
Происхождение и развитие хлоропластов изучено еще очень мало и единой точки зрения по этому вопросу пока не существует. Известно, что в молодых, эмбриональных клетках дифференцированных хлоропластов нет. Вместо них имеются так называемые пропластиды . Это очень мелкие (доли микрона) тельца, находящиеся на грани разрешающей способности светового микроскопа. Первоначально они имеют амебовидную форму (несут лопасти), отграничены от цитоплазмы двойной мембраной и не содержат ни внутренних мембран, ни хлорофилла. Внутренние мембраны, образующие ламеллы, развиваются позднее. Имеется несколько гипотез о дальнейшем развитии пропластид. Согласно одной из них, в прозрачной строме пропластиды сначала образуются скопления мельчайших пузырьков, расположенных в правильном порядке наподобие кристаллической решетки. Это скопление пузырьков, каждый из которых одевается собственной мембраной, называется первичной граной . По периферии первичной граны возникают ламеллы, которые распространяются во все стороны от нее. В дальнейшем на них образуются все ламеллярные структуры хлоропласта, в том числе и граны. На свету в них наблюдается отложение пигментов и, прежде всего, хлорофилла.
По другой гипотезе, ламеллы первоначально образуются как складки внутренней мембраны оболочки пропластиды, а не из пузырьков. При этом сначала возникает структура, подобная митохондрии.
Таким образом, эти гипотезы исходят из принципа непрерывности пластид и отрицают их происхождение из других органоидов протопласта и, прежде всего, из цитоплазмы. Однако другие ученые считают, что митохондрии и пластиды по происхождению тесно связаны между собой. Например, удалось показать возникновение митохондрий из зрелых хлоропластов путем «почкования». Впоследствии эти митохондрии могли снова объединяться с хлоропластами. Но все эти гипотезы не получили еще достаточного обоснования, и вопрос о происхождении пластид еще ждет своего решения.
Кроме возникновения из пропластид, хлоропласты могут размножаться путем простого деления. При этом из взрослого хлоропласта образуются две дочерние пластиды, часто неравных размеров. Электронномикроскопическая картина такого деления до сих пор не изучена.
Структура хлоропласта не остается постоянной, она закономерно изменяется в процессе роста клетки. Изменение структуры хлоропластов с возрастом листьев заметно даже в световой микроскоп. Так, молодым листьям обычно соответствует тонкогранулярная структура, листьям среднего возраста - крупногранулярная структура. В стареющих листьях происходит нарушение структуры и деградация хлоропластов.
Хлоропласты - довольно нежные непрочные органоиды. При помещении клетки в дистиллированную воду или гипотонический солевой раствор они быстро разбухают, на их поверхности образуются пузыревидные вздутия, а затем они расплываются. Электронномикроскопические исследования показали, что разбухание происходит в строме, а не в ламеллах. При повреждении клетки хлоропласты, рассматриваемые в обычный микроскоп, сначала становятся грубогранулярными, вздуваются, приобретают пенистый вид и, наконец, гранулярность исчезает. Патологические изменения происходят в хлоропластах листьев и при недостатке в почве элементов минерального питания. Однако хлоропласты некоторых клеток могут обнаруживать и высокую стойкость. Так, у деревьев зеленый цвет коры обусловлен наличием слоя клеток с хлоропластами. Эти хлоропласты прекрасно переносят низкие температуры и переходят в активное состояние, обнаруживаемое по сильному позеленению коры, например, у осины, очень рано весной, когда ночью еще бывают сильные морозы. Низкие зимние температуры переносят также хлоропласты листьев (хвои) наших вечнозеленых хвойных деревьев. При этом, как показали электронномикроскопические исследования, они сохраняют свою сложную внутреннюю организацию.
Лейкопласты
Это мелкие бесцветные пластиды. В световом микроскопе их часто трудно обнаружить, так как они бесцветны и обладают тем же коэффициентом преломления, что и цитоплазма. Обнаружить их можно лишь в случае накопления внутри них крупных включений. Это очень нежные органоиды и при приготовлении срезов живого материала разрушаются даже более легко, чем хлоропласты. Они встречаются во взрослых клетках, скрытых от действия солнечного света: в корнях, корневищах, клубнях (картофель), семенах, сердцевине стеблей, а также в клетках, подвергающихся сильному освещению (клетки кожицы). Часто лейкопласты собираются вокруг ядра, окружая его иногда со всех сторон. Форма лейкопластов очень непостоянна, чаще всего это шаровидные, яйцевидные или веретеновидные образования.

Лейкопласты - органоиды, связанные с образованием запасных питательных веществ - крахмала, белков и жиров. Деятельность лейкопластов специализирована: одни из них накапливают преимущественно крахмал (амилопласты), другие - белки (протеопласты, называемые также алейронопластами), третьи - масла (олеопласты). Лейкопласты клеток кожицы листьев и стеблей нельзя отнести ни к одному из этих типов, так как функция их еще не выяснена.
Амилопласты накапливают крахмал в виде так называемых крахмальных зерен. Это преобладающий тип лейкопластов. Структура амилопластов и механизм образования крахмала с трудом поддаются изучению в световом микроскопе, а в электронном микроскопе исследованы еще слабо. Как предполагают, они образуются из пропластид, но в отличие от хлоропластов развитие их структуры не идет далеко, а задерживается на довольно ранней стадии - стадии незрелой, слаболамеллярной пластиды. Снаружи амилопласты отграничены двухмембранной оболочкой. Внутри пластида заполнена тонкогранулярной стромой. Образованию крахмальных зерен в амилопластах предшествует развитие мельчайших пузырьков, которые сливаются, уплощаются, ограничивая мембраной участок стромы в центре пластиды. Этот участок, получивший название образовательного центра , становится более светлым, напоминая вакуолю. В образовательном центре и начинается отложение крахмала. Когда будущее крахмальное зерно начинает увеличиваться в размерах, мембраны, отграничивающие образовательный центр, исчезают, в дальнейшем рост зерна идет вне связи с ними. При отложении крахмала оболочка амилопласта и строма могут сильно растягиваться, в результате чего размер амилопласта сильно увеличивается за счет растущего крахмального зерна. Крахмальное зерно заполняет затем почти всю полость амилопласта, оттесняя его живое содержимое на периферию в виде тончайшей пленки на поверхности зерна. Во многих случаях крахмальное зерно может достигать таких размеров, что амилопласт разрывается и сохраняется только на одной стороне крахмального зерна. В этом случае новые порции крахмала могут синтезироваться лишь в тех участках, где крахмальное зерно остается в контакте с мембранами и стромой амилопласта.
Развиваясь из пропластид, амилопласты при определенных условиях могут превращаться в пластиды других типов. Если, например, поместить корень ячменя на свет, то можно видеть, что некоторые лейкопласты увеличиваются в объеме и превращаются в хлоропласты, сходные с теми, которые образуются в листьях. Если такой корень вновь лишить света, то эти хлоропласты уменьшаются в размерах и теряют свой хлорофилл, но не превращаются опять в лейкопласты, а вырабатывают глобулы (шарики) каротина, становясь таким образом хромопластами. Олеопласты , т. е. лейкопласты, образующие главным образом масла, встречаются значительно реже, чем амилопласты (например, в клетках листьев некоторых однодольных). Они обычно представляют собой продукт старения хлоропластов, которые теряют хлорофилл. При этом в строме пластиды возникают мельчайшие глобулы масла. Затем пластидная оболочка разрушается, и содержимое соседних пластид сливается, образуя более крупные жировые капли. Иногда в таких пластидах одновременно накапливается и крахмал.
Синтез запасного белка - протеина - может осуществляться в третьем типе лейкопластов - протеопластах . Белок в виде кристаллов и зерен образуется в семенах многих растений, особенно тех, которые содержат еще и много масла (например, семена клещевины). Протеопласты, подобно амилопластам, возникают из пропластид. Их развитие также задерживается на стадии незрелой ламеллярной пластиды. В строме протеопласта запасной белок первоначально накапливается в виде фибрилл, которые затем объединяются в более крупные пучки. Далее оболочка и строма пластиды разрушаются, а пучки белковых фибрилл превращаются в подобие мелких вязких вакуолей. Затем вакуоли соседних пластид сливаются, часть белка оформляется в виде кристаллоидов.
Таким образом, и крахмал, и запасной белок, и капли масла являются инертными включениями, продуктами жизнедеятельности пластид. Причем каждый из них может накапливаться не только в лейкопластах, но также в хлоропластах и хромопластах. Но если крахмал образуется в пластидах, то запасные белки и жиры очень часто могут иметь и внепластидное происхождение, возникая, вероятно, непосредственно в цитоплазме и независимо от пластид. Структурные процессы, происходящие при этом, до сих пор изучены слабо.
Хромопласты
Хромопласты представляют собой пластиды желтого или оранжевого и даже красного цвета. Они встречаются в клетках многих лепестков (одуванчик, лютик, калужница), зрелых плодов (томаты, шиповник, рябина, тыква, арбуз, апельсин), корнеплодов (морковь, кормовая свекла). Яркий цвет этих органов обусловлен желтыми и оранжевыми пигментами - каротиноидами, сосредоточенными в хромопластах. Эти пигменты характерны и для хлоропластов, но там они маскируются хлорофиллом. Они не растворимы в воде, но растворимы в жирах.
В отличие от хлоропластов форма хромопластов очень изменчива и определяется их происхождением и состоянием в них пигментов, а также систематическим положением образующего их растения. В зависимости от состояния каротиноидов различают хромопласты трех типов:
- хромопласты, у которых каротиноиды откладываются в форме мелких, но видимых в световой микроскоп ни с чем не связанных кристаллов (хромопласты моркови);
- хромопласты, у которых каротиноиды растворены в субмикроскопических липоидных глобулах (лепестки лютика и алоэ);
- хромопласты, каротиноиды которых собраны в пучки, состоящие из субмикроскопических нитей и связанные с фибриллами белка (красный перец, томаты, мандарин).
В отличие от хлоропластов и лейкопластов хромопласты редко возникают непосредственно из пропластид, а обычно представляют собой результат дегенерации хлоропластов. Исключение составляют хромопласты моркови, которые возникают не из хлоропластов, а из лейкопластов или непосредственно из пропластид. Части корнеплода, не погруженные в почву и развивающиеся на свету, обычно зеленеют. Это происходит не в результате превращения хромопластов в хлоропласты, а вследствие образования хлоропластов из пропластид или лейкопластов. Хромопласты вообще не могут превращаться в другие типы пластид. Чаще всего хромопласты образуются при разрушении хлоропластов, когда последние вступают в необратимую фазу развития. Таково происхождение хромопластов 2-го и 3-го типов. При этом в хлоропластах увеличивается содержание жиров и каротиноидов, которые собираются в строме пластиды в виде субмикроскопических глобул, ламеллярные структуры исчезают, а хлорофилл разрушается. Глобулы пигмента растут, а объем стромы уменьшается, в результате глобулы могут заполнить большую часть пластиды. Округлая форма «материнского» хлоропласта при этом сохраняется. Подобный процесс деградации хлоропластов происходит, вероятно, и при осеннем пожелтении листьев и при созревании плодов. Хлорофилл в желтеющих листьях разрушается и перестает маскировать каротиноиды, которые резко выступают и обусловливают желтую окраску листьев.
В корнеплодах моркови хромопласты возникают из лейкопластов, вначале крахмалоносных, при этом в строме пластиды накапливаются каротиноиды, которые позже кристаллизуются. Крахмал исчезает по мере того, как растет концентрация каротина, пластидная масса уменьшается и ее становится трудно обнаружить. Выкристаллизовавшийся пигмент составляет преобладающую по объему часть хромопласта, поэтому форма хромопласта в конечном счете определяется формой кристаллизующегося пигмента и бывает обычно неправильной: зубчатой, серповидной, игольчатой или пластинчатой. Пластинки могут иметь очертания треугольника, ромба, параллелограмма и т. д.

На рисунке изображена одна из клеток арбуза с малиновой мякотью при рассматривании в световой микроскоп. В клетке видна цитоплазма, состоящая из тонких нитей, растянутых в различных направлениях. В более массивных тяжах цитоплазмы расположены игольчатые кристаллы пигмента хромопластов. Наибольшее скопление кристаллов наблюдается около ядра. У другого сорта арбуза с мякотью карминного цвета пигмент хромопластов кристаллизуется не только в виде игольчатых кристаллов, но и коротких призмочек различного размера.
Значение хромопластов в обмене веществ выяснено очень мало. Как и лейкопласты, они лишены способности к фотосинтезу, так как не содержат хлорофилла. Косвенное значение хромопластов состоит в том, что они обусловливают яркую окраску цветков и плодов, привлекающую насекомых для перекрестного опыления и других животных - для распространения плодов.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Пластиды - это органоиды клеток растений и некоторых фотосинтезирующих простейших. У животных и грибов пластид нет.
Пластиды делятся на несколько типов. Наиболее важный и известный - хлоропласт, содержащий зеленый пигмент хлорофилл, который обеспечивает процесс фотосинтеза.
Другими видами пластид являются разноцветные хромопласты и бесцветные лейкопласты. Также выделяют амилопласты, липидопласты, протеинопласты, которые часто считают разновидностями лейкопластов.
Виды пластид: хлоропласты, хромопласты, лейкопласты
Все виды пластид связаны между собой общим происхождением или возможным взаимопревращением. Пластиды развиваются из пропластид – более мелких органоидов меристематических клеток.

Строение пластид
Большинство пластид относится к двумембранным органоидам, у них есть внешняя и внутренняя мембраны. Однако встречаются организмы, чьи пластиды имеют четыре мембраны, что связано с особенностями их происхождения.
Во многих пластидах, особенно в хлоропластах, хорошо развита внутренняя мембранная система, формирующая такие структуры как тилакоиды, граны (стопки тилакоидов), ламелы – удлиненные тилакоиды, соединяющие соседние граны. Внутренне содержимое пластид обычно называют стромой. В ней помимо прочего находятся крахмальные зерна.
Считается, что в процессе эволюции пластиды появились аналогично митохондриям - путем внедрения в клетку-хозяина другой прокариотической клетки, способной в данном случае к фотосинтезу. Поэтому пластиды считают полуавтономными органеллами. Они могут делиться независимо от делений клетки, у них есть собственная ДНК, РНК, рибосомы прокариотического типа, т. е. собственный белоксинтезирующий аппарат. Это не значит, что в пластиды не поступают белки и РНК из цитоплазмы. Часть генов, управляющей их функционированием, находится как раз в ядре.
Функции пластид
Функции пластид зависят от их типа. Хлоропласты выполняют фотосинтезирующую функцию. В лейкопластах накапливаются запасные питательные вещества: крахмал в амилопластах, жиры в элайопластах (липидопластах), белки в протеинопластах.
Хромопласты, за счет содержащихся в них пигментов-каротиноидов, окрашивают различные части растений – цветки, плоды, корнеплоды, осенние листья и др. Яркий окрас часто служит своеобразным сигналом для животных-опылителей и распространителей плодов и семян.
В дегенерирующих зеленых частях растений хлоропласты превращаются в хромопласты. Пигмент хлорофилл разрушается, поэтому остальные пигменты, несмотря на малое количество, становятся в пластидах заметными и окрашивают туже листву в желто-красные оттенки.
На заре развития жизни на Земле все клеточные формы были представлены бактериями. Они всасывали органические вещества, растворённые в первичном океане, через поверхность тела.
Со временем некоторые бактерии приспособились производить органические вещества из неорганических. Для этого они использовали энергию солнечного света. Возникла первая экологическая система, в которой эти организмы были производителями. В результате этого в атмосфере Земли появился кислород, выделяемый этими организмами. С его помощью можно из той же самой пищи получить гораздо больше энергии, а добавочную энергию использовать на усложнение строения тела: разделение тела на части.
Одно из важных достижений жизни — разделение ядра и цитоплазмы. В ядре находится наследственная информация. Специальная мембрана вокруг ядра позволила защитить от случайных повреждений. По мере необходимости цитоплазма получает из ядра команды, направляющие жизнедеятельность и развитие клетки.
Организмы, у которых ядро отделено от цитоплазмы, образовали надцарство ядерных (к ним относятся — растения, грибы, животные).
Таким образом, клетка — основа организации растений и животных — возникла и развилась в ходе биологической эволюции.
Даже не вооружённым глазом, а ещё лучше под лупой можно видеть, что мякоть зрелого арбуза состоит из очень мелких крупинок, или зёрнышек. Это клетки — мельчайшие «кирпичики», из которых состоят тела всех живых организмов, в том числе и растительных.
Жизнь растения осуществляется соединённой деятельностью его клеток, создающих единое целое. При многоклеточности частей растения существует физиологическое разграничение их функций, специализация различных клеток в зависимости от местоположения их в теле растения.
Растительная клетка отличается от животной тем, что имеет плотную оболочку, покрывающую внутреннее содержимое со всех сторон. Клетка не является плоской (как её принято изображать), она скорей всего похожа на очень маленький пузырёк, наполненный слизистым содержимым.
Строение и функции растительной клетки
Рассмотрим клетку как структурно-функциональную единицу организма. Снаружи клетка покрыта плотной клеточной стенкой, в которой имеются более тонкие участки — поры. Под ней находится очень тонкая плёнка — мембрана, покрывающая содержимое клетки — цитоплазму. В цитоплазме есть полости — вакуоли, заполненные клеточным соком. В центре клетки или около клеточной стенки расположено плотное тельце — ядро с ядрышком. От цитоплазмы ядро отделено ядерной оболочкой. По всей цитоплазме распределены мелкие тельца — пластиды.
Строение растительной клетки
Строение и функции органоидов растительной клетки
| Органоид | Рисунок | Описание | Функция | Особенности |
Клеточная стенка или плазматическая мембрана | Бесцветная, прозрачная и очень прочная | Пропускает в клетку и выпускает из клетки вещества. | Клеточная мембрана полупроницаемая |
|
Цитоплазма | Густое тягучее вещество | В ней располагаются все другие части клетки | Находится в постоянном движении |
|
Ядро (важная часть клетки) | Округлое или овальное | Обеспечивает передачу наследственных свойств дочерним клеткам при делении | Центральная часть клетки |
|
Сферической или неправильной формы | Принимает участие в синтезе белка | |||
 | Резервуар, отделённый от цитоплазмы мембраной. Содержит клеточный сок | Накапливаются запасные питательные вещества и продукты жизнедеятельности ненужные клетке. | По мере роста клетки мелкие вакуоли сливаются в одну большую (центральную) вакуоль |
|
Пластиды | Хлоропласты | Используют световую энергию солнца и создают органические из неорганических | Форма дисков, отграниченных от цитоплазмы двойной мембраной |
|
Хромопласты | Образуются в результате накопления каротиноидов | Жёлтые, оранжевые или бурые |
||
 | Лейкопласты | Бесцветные пластиды | ||
Ядерная оболочка | Состоит из двух мембран (наружная и внутренняя) с порами | Отграничивает ядро от цитоплазмы | Даёт возможность осуществляться обмену между ядром и цитоплазмой |
Живая часть клетки — это ограниченная мембраной, упорядоченная, структурированная система биополимеров и внутренних мембранных структур, участвующих в совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Важной особенностью является то, что в клетке нет открытых мембран со свободными концами. Клеточные мембраны всегда ограничивают полости или участки, закрывая их со всех сторон.

Современная обобщенная схема растительной клетки
Плазмалемма (наружная клеточная мембрана) — ультрамикроскопическая плёнка толщиной 7,5 нм., состоящая из белков, фосфолипидов и воды. Это очень эластичная плёнка, хорошо смачивающаяся водой и быстро восстанавливающая целостность после повреждения. Имеет универсальное строение, т.е.типичное для всех биологических мембран. У растительных клеток снаружи от клеточной мембраны находится прочная, создающая внешнюю опору и поддерживающая форму клетки клеточная стенка. Она состоит из клетчатки (целлюлозы) — нерастворимого в воде полисахарида.
Плазмодесмы растительной клетки, представляют собой субмикроскопические канальцы, пронизывающие оболочки и выстланные плазматической мембраной, которая таким образом переходит из одной клетки в другую, не прерываясь. С их помощью происходит межклеточная циркуляция растворов, содержащих органические питательные вещества. По ним же идёт передача биопотенциалов и другой информации.
Порами называют отверстия во вторичной оболочке, где клетки разделяют лишь первичная оболочка и срединная пластинка. Участки первичной оболочки и срединную пластинку, разделяющие соседствующие поры смежных клеток, называют поровой мембраной или замыкающей пленкой поры. Замыкающую пленку поры пронизывают плазмодесменные канальцы, но сквозного отверстия в порах обычно не образуется. Поры облегчают транспорт воды и растворенных веществ от клетки к клетке. В стенках соседних клеток, как правило, одна против другой, образуются поры.
Клеточная оболочка имеет хорошо выраженную, относительно толстую оболочку полисахаридной природы. Оболочка растительной клетки продукт деятельности цитоплазмы. В её образовании активное участие принимает аппарат Гольджи и эндоплазматическая сеть.
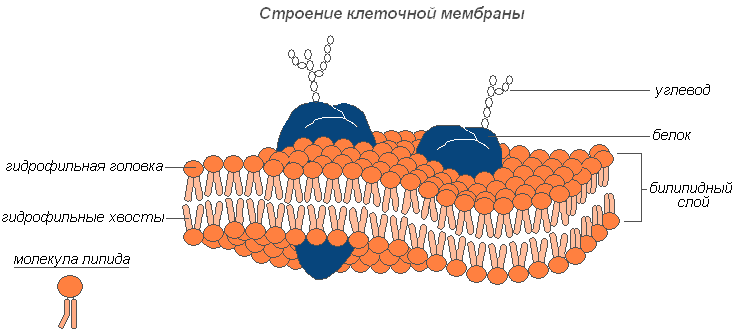
Строение клеточной мембраны
Основу цитоплазмы составляет ее матрикс, или гиалоплазма, — сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель. Важнейшая роль гиалоплазмы заключается в объединении всех клеточных структур в единую систему и обеспечении взаимодействия между ними в процессах клеточного метаболизма.
Гиалоплазма (или матрикс цитоплазмы) составляет внутреннюю среду клетки. Состоит из воды и различных биополимеров (белков, нуклеиновых кислот, полисахаридов, липидов), из которых основную часть составляют белки различной химической и функциональной специфичности. В гиалоплазме содержатся также аминокислоты, моносахара, нуклеотиды и другие низкомолекулярные вещества.
Биополимеры образуют с водой коллоидную среду, которая в зависимости от условий может быть плотной (в форме геля) или более жидкой (в форме золя), как во всей цитоплазме, так и в отдельных ее участках. В гиалоплазме локализуются и взаимодействуют между собой и средой гиалоплазмы различные органеллы и включения. При этом расположение их чаще всего специфично для определенных типов клеток. Через билипидную мембрану гиалоплазма взаимодействует с внеклеточной средой. Следовательно, гиалоплазма является динамической средой и играет важную роль в функционировании отдельных органелл и жизнедеятельности клеток в целом.
Цитоплазматические образования – органеллы
Органеллы (органоиды) — структурные компоненты цитоплазмы. Они имеют определённую форму и размеры, являются обязательными цитоплазматическими структурами клетки. При их отсутствии или повреждении клетка обычно теряет способность к дальнейшему существованию. Многие из органоидов способны к делению и самовоспроизведению. Размеры их настолько малы, что их можно видеть только в электронный микроскоп.
Ядро
Ядро — самая заметная и обычно самая крупная органелла клетки. Оно впервые было подробно исследовано Робертом Броуном в 1831 году. Ядро обеспечивает важнейшие метаболические и генетические функции клетки. По форме оно достаточно изменчиво: может быть шаровидным, овальным, лопастным, линзовидным.
Ядро играет значительную роль в жизни клетки. Клетка, из которой удалили ядро, не выделяет более оболочку, перестаёт расти и синтезировать вещества. В ней усиливаются продукты распада и разрушения, вследствие этого она быстро погибает. Образование нового ядра из цитоплазмы не происходит. Новые ядра образуются только делением или дроблением старого.
Внутреннее содержимое ядра составляет кариолимфа (ядерный сок), заполняющая пространство между структурами ядра. В нём находится одно или несколько ядрышек, а также значительное количество молекул ДНК, соединённых со специфическими белками — гистонами.

Строение ядра
Ядрышко
Ядрышко — как и цитоплазма, содержит преимущественно РНК и специфические белки. Важнейшая его функция заключается в том, что в нём происходит формирование рибосом, которые осуществляют синтез белков в клетке.
Аппарат Гольджи
Аппарат Гольджи — органоид, имеющий универсальное распространение во всех разновидностях эукариотических клеток. Представляет собой многоярусную систему плоских мембранных мешочков, которые по периферии утолщаются и образуют пузырчатые отростки. Он чаще всего расположен вблизи ядра.

Аппарат Гольджи
В состав аппарата Гольджи обязательно входит система мелких пузырьков (везикул), которые отшнуровываются от утолщённых цистерн (диски) и располагаются по периферии этой структуры. Эти пузырьки играют роль внутриклеточной транспортной системы специфических секторных гранул, могут служить источником клеточных лизосом.
Функции аппарата Гольджи состоят также в накоплении, сепарации и выделении за пределы клетки с помощью пузырьков продуктов внутриклеточного синтеза, продуктов распада, токсических веществ. Продукты синтетической деятельности клетки, а также различные вещества, поступающие в клетку из окружающей среды по каналам эндоплазматической сети, транспортируются к аппарату Гольджи, накапливаются в этом органоиде, а затем в виде капелек или зёрен поступают в цитоплазму и либо используются самой клеткой, либо выводятся наружу. В растительных клетках Аппарат Гольджи содержит ферменты синтеза полисахаридов и сам полисахаридный материал, который используется для построения клеточной оболочки. Предполагают, что он участвует в образовании вакуолей. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году.
Лизосомы
Лизосомы представляют собой мелкие пузырьки, ограниченные мембраной основная функция которых — осуществление внутриклеточного пищеварения. Использование лизосомного аппарата происходит при прорастании семени растения (гидролиз запасных питательных веществ).
Строение лизосомы
Микротрубочки
Микротрубочки — мембранные, надмолекулярные структуры, состоящие из белковых глобул, расположенных спиральными или прямолинейными рядами. Микротрубочки выполняют преимущественно механическую (двигательную) функцию, обеспечивая подвижность и сокращаемость органоидов клетки. Располагаясь в цитоплазме, они придают клетке определённую форму и обеспечивают стабильность пространственного расположения органоидов. Микротрубочки способствуют перемещению органоидов в места, которые определяются физиологическими потребностями клетки. Значительное количество этих структур расположено в плазмалемме, вблизи клеточной оболочки, где они участвуют в формировании и ориентации целлюлозных микрофибрилл оболочек растительных клеток.

Строение микротрубочки
Вакуоль
Вакуоль — важнейшая составная часть растительных клеток. Она представляет собой своеобразную полость (резервуар) в массе цитоплазмы, заполненную водным раствором минеральных солей, аминокислот, органических кислот, пигментов, углеводов и отделённую от цитоплазмы вакуолярной мембраной — тонопластом.
Цитоплазма заполняет всю внутреннюю полость только у самых молодых растительных клеток. С ростом клетки существенно изменяется пространственное расположение вначале сплошной массы цитоплазмы: у неё появляются заполненные клеточным соком небольшие вакуоли, и вся масса становится ноздреватой. При дальнейшем росте клетки отдельные вакуоли сливаются, оттесняя к периферии прослойки цитоплазмы, в результате чего в сформированной клетке находится обычно одна большая вакуоль, а цитоплазма со всеми органеллами располагаются около оболочки.
Водорастворимые органические и минеральные соединения вакуолей обусловливают соответствующие осмотические свойства живых клеток. Этот раствор определённой концентрации является своеобразным осмотическим насосом для регулируемого проникновения в клетку и выделения из неё воды, ионов и молекул метаболитов.
В комплексе со слоем цитоплазмы и её мембранами, характеризующимися свойствами полупроницаемости, вакуоль образует эффективную осмотическую систему. Осмотически обусловленными являются такие показатели живых растительных клеток, как осмотический потенциал, сосущая сила и тургорное давление.

Строение вакуоли
Пластиды
Пластиды — самые крупные (после ядра) цитоплазматические органоиды, присущие только клеткам растительных организмов. Они не найдены только у грибов. Пластиды играют важную роль в обмене веществ. Они отделены от цитоплазмы двойной мембранной оболочкой, а некоторые их типы имеют хорошо развитую и упорядоченную систему внутренних мембран. Все пластиды едины по происхождению.
Хлоропласты — наиболее распространённые и наиболее функционально важные пластиды фотоавтотрофных организмов, которые осуществляют фотосинтетические процессы, приводящие в конечном итоге к образованию органических веществ и выделению свободного кислорода. Хлоропласты высших растений имеют сложное внутреннее строение.

Строение хлоропласта
Размеры хлоропластов у разных растений неодинаковы, но в среднем диаметр их составляет 4-6 мкм. Хлоропласты способны передвигаться под влиянием движения цитоплазмы. Кроме того, под воздействием освещения наблюдается активное передвижение хлоропластов амебовидного типа к источнику света.
Хлорофилл — основное вещество хлоропластов. Благодаря хлорофиллу зелёные растения способны использовать световую энергию.
Лейкопласты (бесцветные пластиды) представляют собой чётко обозначенные тельца цитоплазмы. Размеры их несколько меньше, чем размеры хлоропластов. Более и однообразна и их форма, приближающая к сферической.

Строение лейкопласта
Встречаются в клетках эпидермиса, клубнях, корневищах. При освещении очень быстро превращаются в хлоропласты с соответствующим изменением внутренней структуры. Лейкопласты содержат ферменты, с помощью которых из излишков глюкозы, образованной в процессе фотосинтеза, в них синтезируется крахмал, основная масса которого откладывается в запасающих тканях или органах (клубнях, корневищах, семенах) в виде крахмальных зёрен. У некоторых растений в лейкопластах откладываются жиры. Резервная функция лейкопластов изредка проявляется в образовании запасных белков в форме кристаллов или аморфных включений.
Хромопласты в большинстве случаев являются производными хлоропластов, изредка — лейкопластов.

Строение хромопласта
Созревание плодов шиповника, перца, помидоров сопровождается превращением хлоро- или лейкопластов клеток мякоти в каратиноидопласты. Последние содержат преимущественно жёлтые пластидные пигменты — каратиноиды, которые при созревании интенсивно синтезируются в них, образуя окрашенные липидные капли, твёрдые глобулы или кристаллы. Хлорофилл при этом разрушается.
Митохондрии
Митохондрии — органеллы, характерные для большинства клеток растений. Имеют изменчивую форму палочек, зёрнышек, нитей. Открыты в 1894 году Р. Альтманом с помощью светового микроскопа, а внутреннее строение было изучено позднее с помощью электронного.

Строение митохондрии
Митохондрии имеют двухмембранное строение. Внешняя мембрана гладкая, внутренняя образует различной формы выросты — трубочки в растительных клетках. Пространство внутри митохондрии заполнено полужидким содержимым (матриксом), куда входят ферменты, белки, липиды, соли кальция и магния, витамины, а также РНК, ДНК и рибосомы. Ферментативный комплекс митохондрий ускоряет работу сложного и взаимосвязанного механизма биохимических реакций, в результате которых образуется АТФ. В этих органеллах осуществляется обеспечение клеток энергией — преобразование энергии химических связей питательных веществ в макроэргиеские связи АТФ в процессе клеточного дыхания. Именно в митохондриях происходит ферментативное расщепление углеводов, жирных кислот, аминокислот с освобождением энергии и последующим превращением её в энергию АТФ. Накопленная энергия расходуется на ростовые процессы, на новые синтезы и т. д. Митохондрии размножаются делением и живут около 10 дней, после чего подвергаются разрушению.
Эндоплазматическая сеть
Эндоплазматическая сеть — сеть каналов, трубочек, пузырьков, цистерн, расположенных внутри цитоплазмы. Открыта в 1945 году английским учёным К. Портером, представляет собой систему мембран, имеющих ультрамикроскопическое строение.

Строение эндоплазматической сети
Вся сеть объединена в единое целое с наружной клеточной мембраной ядерной оболочки. Различают ЭПС гладкую и шероховатую, несущую на себе рибосомы. На мембранах гладкой ЭПС находятся ферментные системы, участвующие в жировом и углеводном обмене. Этот тип мембран преобладает в клетках семян, богатых запасными веществами (белками, углеводами, маслами), рибосомы прикрепляются к мембране гранулярной ЭПС, и во время синтеза белковой молекулы полипептидная цепочка с рибосомами погружается в канал ЭПС. Функции эндоплазматической сети очень разнообразны: транспорт веществ как внутри клетки, так и между соседними клетками; разделение клетки на отдельные секции, в которых одновременно проходят различные физиологические процессы и химические реакции.
Рибосомы
Рибосомы — немембранные клеточные органоиды. Каждая рибосома состоит из двух не одинаковых по размеру частичек и может делиться на два фрагмента, которые продолжают сохранять способность синтезировать белок после объединения в целую рибосому.

Строение рибосомы
Рибосомы синтезируются в ядре, затем покидают его, переходя в цитоплазму, где прикрепляются к наружной поверхности мембран эндоплазматической сети или располагаются свободно. В зависимости от типа синтезируемого белка рибосомы могут функционировать по одиночке или объединяться в комплексы — полирибосомы.