Kuidas stomata välja näeb? Peamiste ökoloogiliste taimerühmade omadused. Lehtede vananemine ja lehtede langemine
RINGAD. RINGASTE KLASSIFIKATSIOON.
Kõrgemate taimede organiseerimine põhineb rakkude spetsialiseerumise printsiibil, mis seisneb selles, et iga keharakk ei täida kõiki talle omaseid funktsioone, vaid ainult mõnda, kuid täielikumalt ja täiuslikumalt.
Kangad- stabiilsed, looduslikult korduvad rakukompleksid, mis on päritolult, struktuurilt sarnased ja kohandatud täitma üht või mitut funktsiooni.
Kangastel on erinevaid klassifikatsioone, kuid need on kõik üsna meelevaldsed.
Sõltuvalt põhifunktsioonist eristatakse mitut taimekudede rühma.
1. õppekangad, või meristeemid,- on võime jagada ja moodustada kõiki teisi kudesid.
2. Kattavad kuded:
Esmane;
Teisene;
Tertsiaarne.
3. Peamised kangad- moodustavad suurema osa taimekehast. Eristatakse järgmisi peamisi kangaid:
Assimilatsioon (klorofülli kandev);
Kaubahoidjad;
Õhus leviv (aerenhüüm);
Põhjaveekihid.
4. Mehaanilised kangad(toetav, skelett):
kollenhüüm;
Sklerenhüüm.
5. Juhtivad kangad:
Ksüleem (puit) on tõusev kude;
Floem (phloem) on laskuva voolu kude.
6. Ekskretoorsed kuded:
Väline:
Näärmekarvad;
Hüdatoodid – vesistoomid;
Päikeselinnud;
Sisemine:
Eeterlike õlide, vaikude, parkainetega eritavad rakud;
Mitmerakulised eritiste anumad, piimhappelised.
Rakkude jagunemisvõime põhjal eristatakse kahte tüüpi kudesid: hariv, või meristeemid, Ja püsiv- integumentaarne, eritav, aluseline, mehaaniline, juhtiv.
Kangast nimetatakse lihtne, kui kõik selle rakud on kuju ja funktsiooni poolest identsed (parenhüüm, sklerenhüüm, kollenhüüm). Kompleksne kuded koosnevad rakkudest, mis on erineva kuju, struktuuri ja funktsiooniga, kuid on seotud ühise päritoluga (näiteks ksüleem, floeem).
Samuti on olemas kudede klassifikatsioon nende päritolu järgi (ontogeneetiline). Selle klassifikatsiooni järgi eristatakse primaarseid ja sekundaarseid kudesid. Esmasest meristeemist, mis asub võrse ülaosas ja juure tipus, samuti seemne embrüost, esmased konstandid kuded (epidermis, kollenhüüm, sklerenhüüm, assimilatsioonikude, epibleem). Püsivate kudede rakud ei ole võimelised edasiseks jagunemiseks. Spetsiaalse meristeemi rakkudest - prokambia - moodustuvad esmased juhid kuded (primaarne ksüleem, esmane floeem).
Sekundaarsest meristeemist - kambium - moodustuvad teisejärguline koed: sekundaarne ksüleem, sekundaarne floeem; alates fellogeen moodustuvad kork, phelloderm ja läätsed, mis tekivad varre ja juure paksenemisel. Sekundaarseid kudesid leidub tavaliselt iluseemnetaimedes ja kaheidulehelistes katteseemnetaimedes. Puittaimedele on omane sekundaarsete kudede – puidu ja võsa – võimas areng.
HARIDUSRINGAD
Õppekangad Tänu oma rakkude pidevale mitootilisele jagunemisele tagavad nad kõikide taimekudede moodustumise, s.t. tegelikult kujundab ta keha. Iga rakk oma arengus läbib kolm etappi: embrüo-, kasvu- ja diferentseerumisfaasi (st rakk omandab teatud funktsiooni). Embrüo diferentseerumisel säilib primaarne meristeem ainult tulevase võrse tipus (kasvukoonuses) ja juure tipus - apikaalne (apikaalne) meristeemid. Iga taime embrüo koosneb meristeemrakkudest.
Meristeemide tsütoloogilised omadused. Tüüpilised tunnused väljenduvad kõige selgemalt apikaalsetes meristeemides. Need meristeemid koosnevad isodiameetrilistest hulktahulistest rakkudest, mida ei eralda rakkudevahelised ruumid. Nende kestad on õhukesed, sisaldavad vähe tselluloosi ja venivad.
Iga raku õõnsus on täidetud tiheda tsütoplasmaga, millel on suhteliselt suur tuum, mis hõivab keskse positsiooni ja jaguneb intensiivselt mitoosi teel. Hüaloplasma sisaldab palju hajusalt hajutatud ribosoome, proplastiide, mitokondreid ja diktüosoome. Vakuoole on vähe ja need on väikesed. Juhtkuded moodustuvad meristeemist, millel on prosenhümaalne kuju ja suured vakuoolid - prokambium ja kambium. Prokambiumi rakud on ristlõikelt hulknurksed, kambiumirakud on ristkülikukujulised.
Rakud, mis säilitavad oma meristemaatilisi omadusi, jätkavad jagunemist, moodustades üha uusi rakke, mida nimetatakse initsiaalid. Mõned tütarrakud diferentseeruvad, muutudes erinevate kudede rakkudeks, neid nimetatakse initsiaalide tuletised. Esialgsed rakud võivad jaguneda määramatult palju kordi ja initsiaalide derivaadid jagunevad üks või mitu korda ja arenevad püsivateks kudedeks.
Päritolu järgi eristatakse primaarseid ja sekundaarseid meristeeme.
Primaarsed meristeemid
Primaarsed meristeemid tekivad otse embrüo meristeemist ja neil on võime jaguneda. Vastavalt oma asukohale taimes võivad primaarsed meristeemid olla apikaalsed (apikaalsed), interkalaarsed (interkalaarne) ja lateraalsed (lateraalsed).
Apikaalsed (apikaalsed) meristeemid- sellised meristeemid, mis paiknevad täiskasvanud taimedes varre- ja juuretippudes ning tagavad kehapikkuse kasvu. Tüvedes, kasvukoonuses, eristatakse kahte meristemaatilist kihti: tunika, millest moodustub kattekude ja primaarse ajukoore perifeerne osa ning korpus, millest primaarse ajukoore sisemine osa ja keskosa. aksiaalne silinder (joon. 2.3).
Riis. 2.3. Varre apikaalsed meristeemid: A- pikisuunaline läbilõige: 1 - kasvukoonus; 2 - lehe primordium; 3 - aksillaarpunga mugul;
Juure otsas on kolm kihti:
1) dermatogeen, millest moodustub esmane kattekihti neelav kude - risoderm;
2) periblema, millest arenevad esmase ajukoore koed;
3) pleroom, moodustades tsentraalse aksiaalse silindri kuded.
Külgmised (külgmised) meristeemid päritolu järgi võivad need olla primaarsed ja sekundaarsed; aksiaalsete elundite ristlõikel näevad nad välja nagu rõngad. Primaarse külgmise meristeemi näide on prokambium ja peritsükkel. Alates prokambia Moodustuvad kambium ja vaskulaarsete kiudude kimpude primaarsed elemendid (primaarne floeem ja primaarne ksüleem), samas kui prokambiumirakud diferentseeruvad otse primaarsete juhtivate kudede rakkudeks.
Külgmised meristeemid paiknevad paralleelselt elundi pinnaga ja tagavad telgorganite paksuse kasvu.
Interkalaarsed meristeemid sagedamini on need primaarsed ja säilivad eraldi aladena aktiivse kasvu tsoonides taime erinevates osades (näiteks lehelehtede alustel, sõlmevahede alustel). Teraviljade sõlmevahede aluses põhjustab selle meristeemi aktiivsus sõlmevahede pikenemist, mis tagab varre pikkuse kasvu.
Sekundaarsed meristeemid
Sekundaarsete meristeemide hulka kuuluvad külgmised ja haavameristeemid.
Külgmised (külgmised) meristeemid esitati kambium Ja fellogeen. Need moodustuvad promeristeemidest (prokambiumist) või püsivatest kudedest nende dediferentseerumise teel. Kambiumirakud jagunevad vaheseintega paralleelselt elundi pinnaga (perikliinselt). Sekundaarse floeemi elemendid arenevad kambiumi poolt väljapoole ladestunud rakkudest ja sekundaarse ksüleemi elemendid arenevad sissepoole ladestunud rakkudest. Kambiumit, mis tekkis püsivatest kudedest dediferentseerumise teel, nimetatakse lisaks Oma struktuuri ja funktsiooni poolest ei erine see kambiumist, mis tekkis promerisemidest. Fellogeen moodustub subepidermaalsetes kihtides (epidermise all) paiknevatest püsivatest kudedest. Periklinaalselt jagades eraldab fellogeen tulevased korkrakud (felleemid) väljapoole ja phellodermi rakud sissepoole. Seega moodustab fellogeen sekundaarse sisekoe - peridermi. Külgmised meristeemid paiknevad paralleelselt elundi pinnaga ja tagavad telgorganite paksuse kasvu.
Haava meristeemid moodustuvad kudede ja elundite kahjustamisel. Kahjustuse ümber elavad rakud diferentseeruvad, hakkavad jagunema ja muunduvad seeläbi sekundaarseks meristeemiks. Nende ülesanne on moodustada parenhüümirakkudest koosnev tihe kaitsekude - kallus. See kude on valkjat või kollakat värvi, selle rakkudel on suured tuumad ja üsna paksud rakuseinad. Kallus tekib pookimise ajal, tagades võsu sulandumise pookealusega, ja pistikute aluses. See võib moodustada juhuslikke juuri ja pungi, seetõttu kasutatakse seda isoleeritud koekultuuride saamiseks.
KATKEKIE
Primaarne sisekude
TO esmane terviklik kudede hulka kuuluvad epidermis, epidermis ise, parastomataalsed rakud, stoomi kaitserakud ja trihhoomid.
Rakuseinas sisalduvad pektiinained ja tselluloos võivad koos moodustumisega alluda lima tekkele lima Ja igemed. Need on polümeersed süsivesikud, mis on seotud pektiinainetega ja neid iseloomustab nende võime veega kokkupuutel tugevalt paisuda. Paistes igemed on kleepuvad ja neid saab niitideks välja tõmmata, samas kui lima on väga udune ja seda ei saa niitideks välja tõmmata. Pektiinliimi leidub Liliaceae, Cruciferae, Malvaceae, Linden ja Rosaceae sugukondade esindajatel, erinevalt tselluloosi limadest, mida esineb palju vähem (näiteks orhideedel).
Stomata
Need on epidermise väga spetsiifilised moodustised, mis koosnevad kahest oakujulisest kaitserakust ja stomataalsest lõhest (nende vahel on teatud tüüpi rakkudevaheline ruum). Neid leidub peamiselt lehtedes, kuid leidub ka varres (joon. 2.6). 
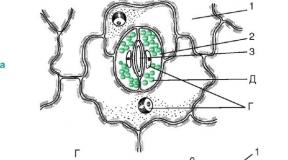
Riis. 2.6. Stomataalne struktuur: a, b- tüümianilehe nahk (pealtvaade ja ristlõige); V- Cereuse (kaktuse perekond) varre koor; 1 - tegelikud epidermise rakud; 2 - stoomi kaitserakud; 3 - kõhulõhe; 4 - õhuõõs; 5 - klorofülli kandva parenhüümi rakud; A - küünenahk; B - küünenaha kiht - kest suberiini ja vahaga; B - seina tselluloosikiht; G - tuum tuumaga; D - kloroplastid
Kaitserakkude seinad on paksenenud ebaühtlaselt: pilu (kõhu) poole suunatud seinad on oluliselt paksemad, võrreldes vahest eemale (selja) suunatud seintega. Vahe võib laieneda ja kahaneda, reguleerides transpiratsiooni ja gaasivahetust. Pilu all on suur hingamisõõs (rakkudevaheline ruum), mida ümbritsevad lehe mesofüllirakud.
Kaitserakke ümbritsevad parastomataalsed rakud, mis koos moodustuvad stomataalne kompleks(joonis 2.7). Eristatakse järgmisi peamisi stomataalsete komplekside tüüpe: 
Riis. 2.7. Stomataalse aparaadi peamised tüübid: 1 - anomotsüütne (kõikides kõrgemates taimedes, välja arvatud korte); 2 - diatsüütiline (sõnajalgadel ja õistaimedel); 3 - paratsüütne (sõnajalgadel, Korteel, õitsev ja rõhuv); 4 - anisotsüütne (ainult õistaimedel); 5 - tetratsüüt (peamiselt üheidulehelistel); 6 - antsüklotsüütne (sõnajalgadel, seemnetaimedel ja õistaimedel)
1) anomotsüütiline(häirega) - kaitserakkudel puuduvad selgelt määratletud parastomataalsed rakud; iseloomulik kõigile kõrgematele taimedele, välja arvatud okaspuud;
2) anisotsüütiline(ebavõrdne rakk) - stoomi kaitserakud on ümbritsetud kolme parastomataalse rakuga, millest üks on teistest palju suurem (või väiksem);
3) paratsüütiline(paralleelrakk) - üks parasomataalne rakk (või rohkem) paikneb paralleelselt kaitserakkudega;
4) diatsüütiline(ristrakk) - kaks parastomataalset rakku asuvad risti kaitserakkudega;
5) tetratsiit(kreeka keelest tetra- neli) - peamiselt üheidulehelised;
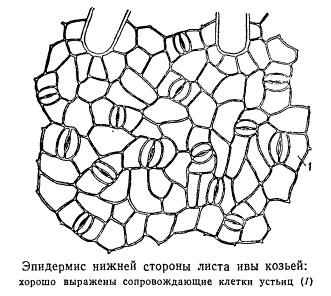
Stoomid paiknevad lehe alumisel küljel, kuid ujulehtedega veetaimedel leidub neid ainult lehe ülemisel küljel. Lehe epidermise rakkude kuju ja stoomi asukoha järgi saab üheidulehelist taime eristada kaheidulehelisest (joon. 2.8). Kaheiduleheliste taimede lehtede tegelikud epidermaalsed rakud on kontuurilt lainelised (joon. 2.9), üheidulehelistel aga piklikud, rombja kujuga. 
Riis. 2.8. Stoomi asukoht epidermis (vaade pinnalt): A-kaheidulehelised taimed: 1 - algustäht; 2 - arbuus; b-ühekojalised: 3 - mais; 4 - iiris
Stoomide tüübid saab jagada vastavalt nende asukoha tasemele epidermise pinna suhtes järgmiselt.
1.7.1. Stoomid, mis asuvad epidermisega samas tasapinnas. Levinuim tüüp ja ravimtaimmaterjalide mikroskoopia kirjelduses seda tavaliselt ei märgita, s.o. see lõik jäetakse välja. Diagnostilised märgid on kas väljaulatuvad või vee all olevad stoomid.
1.7.2. Väljaulatuvad stoomid - epidermise kohal asuvad stoomid. Tavaliselt tuvastatakse mikroskoobi mikroskoobi pööramisel (objektiivi langetamisel) esmalt sellised stoomid ja alles siis ilmuvad epidermise rakud, mistõttu on peaaegu võimatu neid lehe pinnalt fotole jäädvustada. et neid joonisel kujutada. Epidermisega samal tasapinnal võib selliseid stoomi näha põikilõikena, kuid selleks peab lõik läbima stoomi, mida on raske saada, arvestades nende haruldast asukohta lehel. Sellised stoomid on iseloomulikud näiteks karulaugulehtedele.
1.7.3. Sukeldunud stomata - epidermisesse sukeldatud stoomid. Kui vaadeldakse mikroskoobi all mikrokruvi pöörates (läätse langetamise ajal), tuvastatakse kõigepealt selgelt epidermise rakud, seejärel on võimalik selgemalt näha stoomi kontuure. Samuti on keeruline neid pinnalt preparaatide fotodel ja joonistel kuvada. Leitud maikellukese lehtedest, kellalehtedest, eukalüptilehtedest. Mõnikord on süvendid, milles stoomid asuvad, vooderdatud või kaetud karvadega ja neid nimetatakse stomataalsed krüptid.
1.8. Stomataalsete rakkude tüübid
Kirjanduses on kirjeldatud 19 tüüpi, oleme valinud ainult need, mida kasutatakse ravimtaimse tooraine analüüsimisel**.

Riis. 63. Stomataalsete rakkude tüübid. A - läätsekujuline; B - sfääriline; B - korgikujuline; G – abaluud
1.8.1. Läätsekujuline - 2 identset poolkuukujulist lahtrit, mis on paigutatud sümmeetriliselt. Frontaaltasandil on kesta paksenemine peaaegu ühtlane. Lõhe on fusiform (joon. 63, A). Stomataalsete rakkude tüüp on iseloomulik enamikule taimedele.
1.8.2. Sfääriline - kaks ühesugust, tugevalt ringikujuliselt kumerat rakku paiknevad sümmeetriliselt. Frontaaltasandil on kesta paksenemine peaaegu ühtlane. Pilu on ümmargune (joonis 63, B).
1.8.3. Korgikujuline - kahel identsel poolkuukujulisel rakul polaarosades on paksenemised korgi kujul. Lõhe on fusiform (joon. 63, B). Leitud rebaskinnastest.
1.8.4. Scaphoid - stomataalsete rakkude siseseinad on paksenenud. Lõhe on fusiform (joon. 63, D). Täheldatud sajandikrohus ja kellalehtedes.
Stoomi toimimise mehhanismi määravad rakkude osmootsed omadused. Kui lehepind on päikese poolt valgustatud, toimub kaitserakkude kloroplastides aktiivne fotosünteesi protsess. Rakkude küllastumine fotosünteesiproduktide ja suhkrutega toob kaasa kaaliumiioonide aktiivse sisenemise rakkudesse, mille tulemusena suureneb rakumahla kontsentratsioon kaitserakkudes. Parastomataalsete ja kaitserakkude rakumahla kontsentratsioon on erinev. Rakkude osmootsete omaduste tõttu satub parastomataalsetest rakkudest vesi kaitserakkudesse, mis toob kaasa viimaste mahu suurenemise ja turgori järsu suurenemise. Stomataalse lõhe poole jäävate kaitserakkude “kõhu” seinte paksenemine tagab rakuseina ebaühtlase venitamise; kaitserakud omandavad selge oakujulise kuju ja avaneb stomataalne lõhe. Kui fotosünteesi intensiivsus väheneb (näiteks õhtul), väheneb suhkrute moodustumine kaitserakkudes. Kaaliumiioonide sissevool peatub. Rakumahla kontsentratsioon kaitserakkudes on võrreldes parastomataalsete rakkudega vähenenud. Vesi lahkub kaitserakkudest osmoosi teel, alandades nende turgorit; selle tulemusena sulgub stomataalne lõhe öösel.
Epidermise rakud on omavahel tihedalt suletud, tänu millele täidab epidermis mitmeid funktsioone:
Hoiab ära patogeensete organismide tungimise taimesse;
Kaitseb sisekudesid mehaaniliste kahjustuste eest;
Reguleerib gaasivahetust ja transpiratsiooni;
Selle kaudu eraldub vesi ja soolad;
Võib toimida imemiskoena;
võtab osa erinevate ainete sünteesist, ärrituste tajumisest ja lehtede liikumisest.
Trichomes - erineva kuju, struktuuri ja funktsiooniga epidermise rakkude väljakasvud: karvad, soomused, harjased jne. Need jagunevad katvateks ja näärmelisteks. näärmete trihhoomid, erinevalt katetest on neil rakke, mis eritavad eritist. Karvade katmine moodustades taimele villase, vildi või muu katte, peegeldavad nad osa päikesekiirtest ja vähendavad seeläbi transpiratsiooni. Mõnikord leitakse karvu ainult seal, kus stoomid asuvad, näiteks soolehe alumisel küljel. Mõnes taimes suurendavad elusad karvad kogu aurustumispinda, mis aitab kiirendada transpiratsiooni.
Trihhoomi suurused erinevad oluliselt. Pikimad trihhoomid (kuni 5-6 cm) katavad puuvillaseemneid. Kattetrihhoomid on lihtsate ühe- või mitmerakuliste, hargnenud või tähtkujuliste karvade kujul. Kattavad trihhoomid võivad püsida elus pikka aega või kiiresti surra, täites õhuga.
Need erinevad trihhoomidest, mis tekivad ainult epidermise rakkude osalusel. esilekerkijad, mille moodustumisel osalevad ka subepidermaalsete kihtide sügavamal asuvad koed.
Anatoomilised ja diagnostilised tunnused, millel on suurim tähtsus ja suur varieeruvus ravimite tooraine määramisel. Juuksed võivad olla liht- või peakarvad, mis omakorda võivad olla ühe- või mitmerakulised. Mitmerakulised karvad võivad olla üherealised, kaherealised või hargnenud.
LABORITÖÖ nr 5
VEEVAHETUS. LEHT KUI LIIKUMISE ORGAN
Töö eesmärk: taimelehe kui transpiratsiooniorgani olulisemate funktsionaalsete tunnuste uurimine: stoomide ehitus ja arv lehelabal, stoomide avanemise ja sulgumise mehhanism, erinevate ainete mõju stoomi liikumisele.
TRASPIRATSIOONI
Transpiratsiooni bioloogiline tähtsus seisneb esiteks lehe sisetemperatuuri püsivuse tagamises. See saavutatakse soojuse neelamisega vee poolt, kuna see aurustub lehtede poolt. Nimetatakse energiat, mis kulub molekuli ülekandmiseks vedelast faasist gaasilisse olekusse ilma temperatuuri muutmata aurustumissoojus. Soojuse kulutamine vee aurustamisele on vahend lehtede temperatuuri reguleerimiseks ja taimede ülekuumenemise vältimiseks.
Teiseks tagab transpiratsioon, olles ülemise otsa mootor, vee ja mineraaltoitainetega varustamise juurtele. Transpiratsiooni intensiivsuse ning vee ja ioonide juurdevoolu vahel on leitud positiivne korrelatsioon. Kui eemaldate taimelt lehed, peatub vee imendumine juurte poolt. Läbipaistvate lehtede imemisefekti saab kontrollida, kui asetada lõigatud oks veega täidetud pipetti ja langetada elavhõbedatopsi. Mõne aja pärast võite pipetis jälgida elavhõbeda tõusu, mis näitab lehtede märkimisväärset imemisjõudu.
Seega määrab vee juurtesse sisenemise kiiruse transpiratsiooni intensiivsus.
Kolmandaks hoiab transpiratsioon ära turgori ülerõhu tekkimise, mis võib viia taimerakkude hävimiseni.
Neljandaks on transpiratsiooniprotsess tihedalt seotud taimede fotosünteesiga, mida märkisid K. A. Timirjazevi teosed. CO 2 neeldumine taimede lehtedega toimub stoomi kaudu ja see sõltub lehekoe veega küllastumise astmest. Vee ja süsinikdioksiidi assimilatsiooniprotsess on ühtne ja lahutamatu tervik.
Transpiratsiooni kiirust mõistetakse kui lehepinna ühikust ajaühikus aurustunud vee hulka. Tavaliselt on selle indikaatori mõõde mg/dm2 tunnis. Taimede poolt aurustatud vee hulk on üsna suur ja ületab sageli kasvuperioodil sademete hulga. Selle ülejäägi kompenseerivad sügis-talvised sademed. Näiteks üks päevalille- või maisitaim kulutab suve jooksul 200–250 liitrit vett. Nisutaimed 1 hektari suurusel alal aurustavad suve jooksul umbes 2 miljonit liitrit vett, mais - üle 3 miljoni ja kapsas - kuni 8 miljonit liitrit. Ühe kilogrammi taimemassi moodustamisel kulub 300 liitrit. Vesi.
Stomataalset transpiratsiooni reguleerib stoomi avatusaste. Nende struktuur ja levik sõltuvad taimede liigist ja keskkonnaomadustest. Stomateid leidub kõikidel taime maapealsetel osadel, sealhulgas suguelunditel ja isegi tolmuniitidel. Kõige iseloomulikumad stoomid on lehtede jaoks. Sagedamini paiknevad need lehtede alumisel küljel (mesofüütsetes taimedes). Kuid kserofüütidel leidub neid ka lehe ülemisel küljel.
Keskmine stoomide arv 1 mm 2 pindala kohta jääb vahemikku 100 kuni 300. Stoomide suurus ei ületa 20 mikronit pikkust ja 8-15 mikronit laiust. Avatud stoomi kogupindala moodustab 1% lehe pinnast.
On kindlaks tehtud, et väikestel apikaalsetel lehtedel on rohkem stoomi kui suurtel madalamatel. Stoomide esinemissagedus (nende arv pindalaühiku kohta) suureneb, kui liikuda lehe aluselt selle tippu ja taime alt üles. Kuivade kasvukohtade taimedel on neid rohkem, kuid need on väiksema suurusega.
Enamikus mesofüütsetes taimedes paiknevad stoomid epidermise rakkudega samal tasemel ja kserofüütilistes vormides asuvad stoomid epidermise tasemest madalamal ja neid nimetatakse veealuseks. Hügrofüütidel paiknevad kaitserakud mõnikord epidermise kohal. Selliseid stomateid nimetatakse kõrgendatud.
Teatud taimerühmadele on omane üht- või teist tüüpi stomati struktuur, kuigi samas perekonnas võib mõnikord kohata erinevat tüüpi stomaate. Vaatamata stoomide poolt hõivatud märkimisväärsele alale moodustab veeauru difusioon nende kaudu 50–60% vaba pinna aurustumisest. On kindlaks tehtud, et väikeste aukude kaudu levimise kiirus on võrdeline nende ümbermõõduga, mitte nende pindalaga. Seetõttu mõjutab kaitserakkude osaline sulgemine nende perimeetrit vähe ja veeauru difusiooni tase läbi stoomide ei lange väga järsult.
Katse 1. Stoomi liikumise jälgimine mikroskoobi all.
Kogemuse eesmärk: määrake stoomide sõltuvus osmootselt aktiivsetest ainetest.
Materjalid ja varustus: 5% glütseriini lahus, habemenuga, lahkamisnõel, mikroskoop, objektiklaasid ja katteklaasid.
Taimed: lehed (Tradescantia, tulp, hortensia või amaryllis, Kalanchoe).
Gaasivahetust lehtede rakkudevaheliste ruumide ja väliskeskkonna vahel reguleerivad stoomid. Iga stoma koosneb kahest kaitserakust, milles stomataalse lõhega külgnevad seinad on tugevasti paksenenud, samas kui kesta välisosad jäävad õhukeseks. Välis- ja siseseinte ebavõrdne paksus toob kaasa asjaolu, et turgori muutumisel suudavad kaitserakud painduda või sirgendada, avades või sulgedes stomataalse lõhe.
Edusammud: tehakse valitud taime lehe epidermisest lõigud, mis asetatakse 5% glütserooli lahusesse ja hoitakse vähemalt 1 tund.Lõike uuritakse mikroskoobi all ning stomataalse lõhe avanemisaste määratakse määratakse okulaari mikromeetri abil. Tehke 10 mõõtmist, leidke keskmine väärtus ja arvutage keskmise viga. Seejärel viiakse lõigud glütseroolilahusest vette ja stomataalsete pilude mõõtmisi korratakse mikroskoobi all. Tulemused on registreeritud tabelis 1.
Tabel 1
Stomataalse lõhe avanemise aste erinevates keskkondades
|
taim, orel |
Mõõtmise nr. |
Stomataalse lõhe avanemise aste |
|
|
Glütserool | |||
|
taime leht | |||
Harjutus: teha järeldus glütserooli ja vee mõju kohta stoomide avanemisele ja sulgemisele.
Katse 2. Stoomide ja rakkudevaheliste ruumide seisundi määramine Molischi meetodil
Kogemuse eesmärk: määrab välistingimuste mõju stoomi seisundile ja transpiratsiooni intensiivsusele.
Materjalid ja seadmed: ksüleen (tilgutisse), etüülalkohol (tilgutisse); benseen (tilgutisse), pipetid.
Taim: taimede värsked või kuivanud lehed, pimedas olnud taimede lehed.
Lehe rakkudevahelised ruumid on tavaliselt täidetud õhuga, mille tõttu paistab leht valguse käes vaadates matina. Kui teostate infiltratsiooni, st. täites rakkudevahelised ruumid mis tahes vedelikuga, muutuvad lehe vastavad alad läbipaistvaks.
Stoomi seisundi määramine infiltratsioonimeetodi abil põhineb rakumembraane märgavate vedelike võimel tungida kapillaarsuse jõul läbi avatud stomataalsete pilude lähimatesse rakkudevahelistesse ruumidesse, tõrjudes sealt välja õhu, mis on välimuse järgi hästi näha. läbipaistvad laigud lehel. Erinevad vedelikud on võimelised tungima erineval määral avatud stomataalsetesse piludesse: ksüleen tungib kergesti läbi kergelt avatud stoomide, benseen läbi mõõdukalt avatud stoomide ja etüülalkohol suudab tungida ainult läbi laialt avatud stoomide.
See Molischi pakutud meetod on väga lihtne ja üsna rakendatav ka valdkonnas töötamiseks.
Edusammud. Kandke lehe alumisele pinnale eraldi väikesed tilgad benseeni, ksüleeni ja etüülalkoholi. Hoidke lehte horisontaalasendis, kuni tilgad, mis võivad aurustuda või lehe sisse tungida, täielikult kaovad, ja uurige lehte valguse käes.
Uurige erinevates tingimustes (värsked ja närbunud, valgustatud ja varjutatud jne) hoitud lehti. Kontrollige iga kord 2-3 lehte.
tabel 2
Väliste tingimuste mõju kõhuõõne avanemise astmele
Harjutus: Märkige tulemused tabelisse 2, märkides stoomi avatuse astme: lai, keskmine, nõrk. Tehke järeldus välistingimuste mõju kohta stomati liikumisele.
Katse 3. Stoomide seisundi määramine Molotkovski jäljendite abil.
Töö eesmärk: stoomi töö määramine sõltuvalt valgustusest.
Materjalid ja seadmed: värvitu küünelakk, õhuke klaaspulk, pintsetid, mikroskoop, okulaari mikromeeter, esememikromeeter.
Taimed: toataimed, mille lehed kaetakse 2-3 tundi enne tundi valguskindla kattega.
Lehe pinnale kantakse õhuke lakk. Pärast lahusti aurustumist moodustub kile, millele on jäljendatud stoomiga epidermis. Uurides saadud väljatrükke läbi mikroskoobi, saate määrata stoomide arvu ja suuruse ning mõõta stomataalsete pilude laiust. Seda meetodit saab kasutada mitte ainult labori-, vaid ka väliuuringuteks (viimasel juhul säilitatakse väljatrükke kuni määramiseni veega katseklaasides). Nende lehtede uurimiseks, mille stomata asuvad epidermise süvendites (näiteks oleandris), ei saa seda meetodit kasutada, kuna Sellised lehed ei tooda väljatrükke.
Edusammud. Kandke klaaspulgaga lehe alumisele küljele tilk lakilahust ja ajage see kiiresti õhukese kihina laiali. Pärast kuivatamist eemaldage kile pintsettidega, asetage see klaasklaasile ja uurige seda suure suurendusega. Sisestage okulaarne mikromeeter mikroskoopi ja mõõtke stomataalse lõhe laius ja pikkus vähemalt 10 stoomi ulatuses ning arvutage keskmised väärtused.
Määrake silma mikromeetri jaotusväärtus. Selleks asetage mikroskoobi staadiumile mikromeetriline objekt, mille iga jaotus on 0,01 mm või 10 mikronit. Okulaari keerates joondage mõlemad skaalad nii, et nende skaalad oleksid paralleelsed ja üks kattuks teisega. Silma mikromeetri jaotusväärtuse määramine toimub noonuse põhimõttel, s.o. ühendage üks okulaari ja objektiivi mikromeetri skaalajoontest ning leidke järgmine joondus. Leidke sobivad jooned ja määrake, mitu silmamikromeetri A jaotust vastavad kombineeritud punktide vahel paikneva objekti mikromeetri B jaotustele. Silma mikromeetri jaotushind määratakse järgmise valemiga:
Jagamise väärtus = B · 10 µm/A.
Korrutades stomataalsete avade pikkuse ja laiuse, mis on väljendatud silma mikromeetri osades, ühe jaotuse hinnaga, saate leida stomataalsete pilude absoluutmõõtmed. Arvutage stomataalse lõhe pindala ligikaudselt, korrutades pikkuse laiusega.
Uurige sama taime erinevate tasandite lehti, samuti hästi valgustatud ja varjutatud lehti. Kirjutage tulemused tabelisse 3.
Tabel 3
Valgustuse mõju stomataalsete avade suurusele
Harjutus: teha järeldusi kihilisuse ja valgustingimuste mõju kohta stomataalsete avade suurusele.
Kuigi teadlased on juba ammu teadnud vee aurustumisest lehe pinnalt, oli esimene, kes vaatles stomata itaalia loodusteadlane Marcello Malpighi, kes avaldas selle avastuse 1675. aastal oma töös. Anatom plantarum. Kuid ta ei mõistnud nende tegelikku funktsiooni. Samal ajal töötas tema kaasaegne Nehemiah Grew välja hüpoteesi stoomide osalemise kohta taime sisekeskkonna ventilatsioonis ja võrdles neid putukate hingetoruga. Uuringus tehti edusamme 19. sajandil ja siis, aastal 1827, kasutas Šveitsi botaanik Decandolle esimest korda sõna "stoma". Stoomide uurimist viisid sel ajal läbi Hugo von Mohl, kes avastas stoomide avamise põhiprintsiibi, ja Simon Schwendener, kes klassifitseeris stoomid nende struktuuri tüübi järgi.
Mõnda stoomi toimimise aspekti uuritakse ka praegu intensiivselt; Materjaliks on peamiselt Commelina vulgaris ( Commelina communis), aeduba ( Vicia faba), magus mais ( Zea mays) .
Struktuur
Stoomi mõõtmed (pikkus) jäävad vahemikku 0,01-0,06 mm (polüploidsete taimede ja varjus kasvavate lehtede stoomid on suuremad. Suurimad stoomid leiti väljasurnud taimel Zosterophyllum, 0,12 mm (120 µm). Poor koosneb paarist spetsiaalsetest rakkudest, mida nimetatakse kaitserakkudeks ( cellulae claudentes), mis reguleerivad pooride avanemise astet; nende vahel on stomataalne lõhe ( porus stomatalis). Kaitserakkude seinad on paksenenud ebaühtlaselt: pilu (kõhu) poole suunatud seinad on paksemad kui vahest (selja) suunatud seinad. Vahe võib laieneda ja kahaneda, reguleerides transpiratsiooni ja gaasivahetust. Kui vett on vähe, kleepuvad kaitserakud tihedalt üksteise külge ja stomataalne lõhe on suletud. Kui kaitsekambrites on palju vett, avaldab see survet seintele ja õhemad seinad venivad rohkem ning paksemad tõmmatakse sissepoole, tekib kaitsekambrite vahele tühimik. Pilu all on substomataalne (õhu)õõnsus, mida ümbritsevad lehemassi rakud, mille kaudu toimub otsene gaasivahetus. Süsinikdioksiidi (süsinikdioksiidi) ja hapnikku sisaldav õhk tungib nende pooride kaudu lehekoesse ning seda kasutatakse edasi fotosünteesi ja hingamise protsessis. Lehe sisemiste rakkude fotosünteesi käigus toodetud liigne hapnik pääseb nende samade pooride kaudu tagasi keskkonda. Samuti eraldub aurustumisprotsessi käigus läbi pooride veeauru. Epidermaalseid rakke, mis külgnevad tagumiste rakkudega, nimetatakse saatvateks rakkudeks (tagatis-, naaber-, parastomataalsed). Nad osalevad valverakkude liikumises. Kaitse ja sellega kaasnevad rakud moodustavad stomataalse kompleksi (stomatal aparaadi). Stoomi olemasolu või puudumine (stoomi nähtavaid osi nimetatakse stomataalsed jooned) kasutatakse sageli taimede klassifitseerimisel.
Stoomide tüübid
Kaasnevate rakkude arv ja nende asukoht stomataalse lõhe suhtes võimaldavad eristada mitut tüüpi stoomi:
- anomotsüütilised - kaasnevad rakud ei erine teistest epidermise rakkudest, tüüp on väga levinud kõigi kõrgemate taimede rühmade jaoks, välja arvatud okaspuud;
- diatsiit - mida iseloomustavad ainult kaks kaasnevat rakku, mille ühine sein on kaitsekambriga täisnurga all;
- paratsüütilised - kaasnevad rakud paiknevad paralleelselt kaitserakkude ja stomataalse lõhega;
- anisotsüütilised - kaitserakud on ümbritsetud kolme kaasneva rakuga, millest üks on teistest märgatavalt suurem või väiksem, seda tüüpi leidub ainult õistaimedel;
- tetratsütaarne - neli kaasnevat rakku, mis on iseloomulikud üheidulehelistele;
- entsüklotsütaarne - kaasasolevad rakud moodustavad kaitserakkude ümber kitsa ratta;
- aktinotsüüt – mitmed kaasrakud, mis kiirguvad kaitserakkudest;
- peritsüütilised - kaitserakud on ümbritsetud ühe sekundaarse kaasrakuga, stomata ei ole antikliinilise rakuseinaga seotud kaasneva rakuga;
- desmotsüüt - kaitserakud on ümbritsetud ühe kaasrakuga, stomata on sellega ühendatud antikliinilise rakuseinaga;
- polotsüütilised - kaitserakud ei ole täielikult ümbritsetud ühe kaasnevaga: üks või kaks epidermise rakku külgnevad ühe stomataalse poolusega; stomata on kinnitatud ühe kaasneva raku distaalsele küljele, millel on U- või hobuseraua kuju;
- stephanotsüütne - stoomid, mida ümbritsevad neli või enam (tavaliselt viis kuni seitse) halvasti diferentseerunud kaasnevat rakku, moodustades enam-vähem selgelt eristuva roseti;
- laterotsüütne – seda tüüpi stomataalset aparaati peab enamik botaanikuid anomotsüütilise tüübi lihtsaks modifikatsiooniks.
Stomati asukoht
Kaheidulehelistel taimedel on reeglina lehe alumises osas rohkem stoomi kui ülaosas. Seda seletatakse asjaoluga, et horisontaalselt asetseva lehe ülemine osa on reeglina paremini valgustatud ja selles sisalduvate stoomide väiksem arv takistab vee liigset aurustumist. Lehti, mille alumisel küljel asuvad stoomid, nimetatakse hüpostomaatilisteks.
Üheidulehelistel taimedel on stoomi esinemine lehe ülemises ja alumises osas erinev. Väga sageli asetsevad üheidulehelised lehed vertikaalselt, sel juhul võib stoomide arv lehe mõlemal osal olla sama. Selliseid lehti nimetatakse amfistomaatilisteks.
Ujuvatel lehtedel ei ole lehe alumises osas stoomi, et need saaksid läbi küünenaha vett imada. Lehti, mille ülemisel küljel asuvad stoomid, nimetatakse epistomaatilisteks. Veealustel lehtedel pole üldse stomata.
Okaspuu taimede stomata on tavaliselt peidetud sügavale endodermise alla, mis võimaldab talvel ja suvel põua ajal oluliselt vähendada veekulu aurumiseks.
Sammaldel (välja arvatud Anthocerotes) puuduvad tõelised stoomid.
Stoomid erinevad ka oma asukoha poolest epidermise pinna suhtes. Mõned neist asuvad teiste epidermise rakkudega samal tasapinnal, teised on pinnast kõrgemal või alla maetud. Üheiduidulistel, kelle lehed kasvavad valdavalt pikkuses, moodustavad stomata korrapärased paralleelsed read, kaheiduidulistel aga juhuslikult.
Süsinikdioksiid
Kuna süsinikdioksiid on fotosünteesi protsessis üks peamisi reaktiive, on enamikul taimedel stomatid päeva jooksul avatud. Probleem on selles, et õhku sisenedes seguneb see lehelt aurustuva veeauruga ja seetõttu ei saa taim süsihappegaasi juurde, ilma et ta samal ajal vett kaotaks. Paljudel taimedel on kaitse vee aurustumise eest vahaladestuste kujul, mis ummistavad stomata.
Epidermise koesüsteemi kuuluvad stoomid on taime elus eriti olulised. Stoomi struktuur on nii ainulaadne ja nende tähtsus on nii suur, et neid tuleks käsitleda eraldi.
Epidermise koe füsioloogiline tähtsus on kahekordne, suures osas vastuoluline. Ühelt poolt on epidermis struktuurselt kohandatud kaitsma taime kuivamise eest, mida soodustab epidermise rakkude tihe sulgumine, küünenaha teke ja suhteliselt pikad katvad karvad. Kuid teisest küljest peab epidermis läbima veeauru ja erinevate gaaside masse, mis tormavad vastastikku vastassuunas. Gaasi ja auru vahetus võib teatud tingimustel olla väga intensiivne. Taimeorganismis lahendatakse see vastuolu edukalt stoomide abil. Stomat koosneb kahest omapäraselt modifitseeritud epidermaalsest rakust, mis on üksteisega (piki oma pikkuses) vastasotstega ühendatud ja nn. kaitserakud. Nendevahelist rakkudevahelist ruumi nimetatakse kõhulõhe.
Kaitserakke nimetatakse nn, kuna turgori aktiivsete perioodiliste muutuste kaudu muudavad nad oma kuju nii, et stomataalne lõhe kas avaneb või sulgub. Järgmised kaks omadust on nende kõhuliigutuste jaoks väga olulised. Esiteks sisaldavad kaitserakud erinevalt teistest epidermise rakkudest kloroplaste, milles toimub valguse käes fotosüntees ja moodustub suhkur. Suhkru kui osmootselt aktiivse ainena kuhjumine põhjustab kaitserakkude turgorrõhu muutust võrreldes teiste epidermise rakkudega. Teiseks paksenevad kaitserakkude membraanid ebaühtlaselt, mistõttu turgorirõhu muutus põhjustab nende rakkude mahu ebaühtlase muutumise ja sellest tulenevalt ka nende kuju muutumise. Kaitserakkude kuju muutus põhjustab stomataalse lõhe laiuse muutumise. Illustreerime seda järgmise näitega. Joonisel on kujutatud üks kaheiduleheliste taimede stoomi tüüpidest. Stoomi välimine osa koosneb küünenaha poolt moodustatud membraanilistest eenditest, mis on mõnikord ebaolulised ja mõnikord üsna olulised. Need piiravad välispinnast väikest ruumi, mille alumine piir on stomataalne vahe ise, nn. eesõue stomata. Stomataalse pilu taga, sees, on veel üks väike ruum, mida piiravad kaitsekambrite külgseinte väikesed sisemised väljaulatuvad osad, nn. siseõue stomata. Terrass avaneb otse suurde rakkudevahelisse ruumi, mida nimetatakse õhuõõnsus.
Valguses tekib kaitserakkudes suhkur, see tõmbab vett naaberrakkudest, kaitserakkude turgoor suureneb ja nende kesta õhukesed osad venivad rohkem kui paksud. Seetõttu muutuvad stomataalse pilu sisse ulatuvad kumerad eendid tasaseks ja stomat avaneb. Kui näiteks suhkur muutub öösel tärkliseks, siis kaitserakkude turgor langeb, see põhjustab kesta õhukeste osade nõrgenemist, need ulatuvad üksteise poole ja stomata sulgub. Erinevates taimedes võib stomativahe sulgemise ja avamise mehhanism olla erinev. Näiteks kõrrelistel ja tarnadel on kaitserakud laienenud otstega ja keskosas kitsendatud. Rakkude keskmiste osade membraanid on paksenenud, samas kui nende laienenud otsad säilitavad õhukesed tselluloosmembraanid. Turgori suurenemine põhjustab rakkude otste turset ja selle tulemusena liiguvad sirged mediaanosad üksteisest eemale. See viib stoomi avanemiseni.
Stomataalse aparaadi töömehhanismi omadused tulenevad nii kaitserakkude kujust ja struktuurist kui ka stoomiga külgnevate epidermaalsete rakkude osalemisest selles. Kui stoomiga vahetult külgnevad rakud erinevad välimuselt teistest epidermise rakkudest, nimetatakse neid kaasnevad stoomirakud.
Enamasti on kaasnevatel ja järelrakkudel ühine päritolu.
Stoomi kaitserakud on kas epidermise pinnast veidi kõrgemale tõstetud või, vastupidi, langetatud enam-vähem sügavatesse süvenditesse. Sõltuvalt kaitserakkude asendist epidermise pinna üldise taseme suhtes muutub stomataalse lõhe laiuse reguleerimise mehhanism mõnevõrra. Mõnikord muutuvad stoomi kaitserakud lignifitseerituks ja seejärel määrab stoomilõhe avanemise reguleerimise epidermise naaberrakkude aktiivsus. Laienedes ja kahanedes, st muutes oma mahtu, haaravad nad endaga külgnevad kaitserakud. Sageli aga ei sulgu lignified kaitserakkudega stoomid üldse. Sellistel juhtudel toimub gaasi- ja auruvahetuse intensiivsuse reguleerimine erinevalt (nn alustava kuivatamise kaudu). Lignified kaitserakkudega stoomides katab küünenahk sageli üsna paksu kihiga mitte ainult kogu stomataalse lõhe, vaid ulatub isegi õhuõõnsusse, vooderdades selle põhja.
Enamikul taimedel on stoomid lehe mõlemal küljel või ainult alumisel küljel. Kuid on ka taimi, millel stoomid tekivad ainult lehe ülemisel küljel (veepinnal hõljuvatel lehtedel). Lehtedel on reeglina rohkem stomata kui rohelistel vartel.
Stoomide arv erinevate taimede lehtedel on väga erinev. Näiteks varikatuseta broomlehe alumisel küljel on stoomide arv keskmiselt 30 1 mm 2 kohta, samadel tingimustel kasvaval päevalillel umbes 250. Mõnel taimel on kuni 1300 stoomi 1 mm 2 kohta.
Sama taimeliigi isenditel sõltub stoomide tihedus ja suurus tugevalt keskkonnatingimustest. Näiteks täisvalguses kasvanud päevalille lehtedel oli 1 mm 2 lehepinna kohta keskmiselt 220 stoomi ja esimese kõrval kasvanud, kuid kerge varjundiga isendil umbes 140. üks täisvalguses kasvanud taim, tihedus stomata suureneb alumistest lehtedest ülemistele.
Stoomide arv ja suurus ei sõltu tugevalt ainult taime kasvutingimustest, vaid ka eluprotsesside sisesuhetest taimes endas. Need väärtused (koefitsiendid) on kõige tundlikumad reaktiivid iga taime kasvu määravate tegurite kombinatsiooni jaoks. Seetõttu annab erinevates tingimustes kasvatatud taimede lehtede tiheduse ja suuruse määramine aimu iga taime ja selle keskkonnaga seotud suhete olemusest. Kõik meetodid konkreetse elundi anatoomiliste elementide suuruse ja arvu määramiseks kuuluvad kvantitatiivsete anatoomiliste meetodite kategooriasse, mida mõnikord kasutatakse keskkonnauuringutes, aga ka kultuurtaimede sortide iseloomustamiseks, kuna iga kultuurtaime sort on mida iseloomustavad teatud suuruse ja anatoomiliste elementide arvu piirangud pindalaühiku kohta. Kvantitatiivse anatoomia meetodeid saab suure kasuga kasutada nii taimekasvatuses kui ka ökoloogias.
Gaasi- ja auruvahetuseks mõeldud stoomide kõrval on ka stoomid, mille kaudu vesi eraldub mitte auruna, vaid tilk-vedelikus. Mõnikord on sellised stoomid üsna sarnased tavalistega, ainult veidi suuremad ja nende kaitserakkudel puudub liikuvus. Üsna sageli puuduvad sellises täielikult küpses olekus stomatis kaitserakud ja jääb ainult auk, mis juhib vee välja. Nimetatakse stomateid, mis eritavad vedelat vett vesi ja kõik moodustised, mis on seotud tilk-vedeliku vee vabanemisega - hüdatoodid.
Hüdatoodide struktuur on mitmekesine. Mõnel hüdatoodil on vett eemaldava augu all parenhüüm, mis osaleb vee ülekandmisel vett juhtivast süsteemist ja selle vabanemisest elundist; teistes hüdatoodides läheneb vett juhtiv süsteem otse väljalaskeavale. Eriti sageli moodustuvad hüdatoodid erinevate taimede seemikute esimestel lehtedel. Nii eraldavad niiske ja sooja ilmaga teraviljade, herneste ja paljude heinamaade noored lehed tilkhaaval vett. Seda nähtust võib täheldada suve esimesel poolel iga ilusa päeva varahommikul.
Kõige paremini määratletud hüdatoodid asuvad piki lehtede servi. Sageli kannab üks või mitu hüdatoodi iga hambahari, mis lülitab lehtede servad välja.
Lehe kokkupuuteks atmosfääriga on poorid - stomata. Stoma - see on ava (vahe), mida piiravad kaks kaitselahtrit. Stomata leidub kõigis taime maismaaorganites, kuid kõige enam lehtedes. Erinevalt epidermise rakkudest on igal stoomi kaitserakul kloroplast. Mis neis toimub fotosüntees, kuigi väiksema intensiivsusega kui mesofüllirakkudes. Stomata on üks originaalsetest seadmetest, millel on võimalus avada ja sulgeda sõltuvalt kaitserakkude küllastumisest veega. Tavaliselt on hambaavad piiratud kaks valvekambrit, mille seinad on ebaühtlaselt paksenenud. Kaheidulehelistel taimedel on kaitserakud oakujulised ehk poolkuukujulised, samas kui nende üksteisega külgnevad sisemised rakuseinad on paksemad ja välised õhemad. Kaitserakkude protoplastid on seotud ühes tükis perforatsioonid külgnevate ühisseinte aluses. Kui vett on vähe, kleepuvad kaitserakud tihedalt üksteise külge ja stomataalne lõhe on suletud. Kui kaitsekambrites on palju vett, avaldab see survet rakuseintele ja õhemad seinad venivad rohkem välja ning paksemad tõmbuvad sissepoole ning kaitsekambrite vahele tekib tühimik.
Viimasel ajal on tõestatud, et suur tähtsus on ka stoomide asukohal. tselluloosi mikrofibrillid. Kui tavaliselt on leherakkudes tselluloosfibrillid pikuti orienteeritud ja selles suunas paksenenud, siis stoomi kaitserakkudes on mikrofibrillid organiseeritud radiaalselt, mis suurendab vastupidavust venitusprotsessile.
Teraviljades kaitserakkude struktuur on mõnevõrra erinev. Neid esindavad kaks piklikku lahtrit, mille otstes on seinad õhemad. Kui see on veega küllastunud, venivad õhemad seinad otstes välja ja suruvad kaitserakud lahku, põhjustades tühimiku tekkimist.
Stomataalsete avade arv varieerub olenevalt taimetüübist 10 kuni 600 1 mm2 lehe kohta. Paljudel taimedel (75% liikidest), sealhulgas enamikul puitunud taimedel, paiknevad stoomid lehe alaküljel. Stomataalsete lõhede läbimõõt on vaid 3-12 mikronit. Stomatid ühendavad lehe siseruume väliskeskkonnaga. Vesi siseneb lehele veenide võrgu kaudu, milles paiknevad vaskulaarsed elemendid. Võimalikud on kolm aurustumisviisi:
- läbi stoomi - stomatal,
- küünenaha - küünenaha,
- läätsede kaudu - läätsekujuline transpiratsioon.
Küünenaha ja stomataalse transpiratsiooni eristamine tehti esmakordselt 1877. aastal.
Taimelehtede hambaaparaadi peamised tüübid.
- anomotsüütne (kõikides kõrgemates taimedes, välja arvatud korte),
- diatsüütiline (sõnajalgadel ja õistaimedel),
- paratsüütne (sõnajalgadel, korteel, õitsev ja rõhuv),
- anisotsüütne (ainult õistaimedel),
- tetratsüüt (peamiselt üheiduidulistel),
- entsüklotsütaarne (sõnajalgadel, seemnetaimedel ja õistaimedel).
Stoomid on epidermise väga spetsiifilised moodustised, mis koosnevad kahest kaitserakust, mille vahel on teatud tüüpi rakkudevaheline ruum või kõhulõhe.
Vahe võib laieneda ja kahaneda, reguleerides transpiratsiooni ja gaasivahetust. Pilu all on hingamis- ehk õhuõõs, mida ümbritsevad lehe viljaliha rakud. Kaitserakkudega külgnevaid epidermise rakke nimetatakse sekundaarne ehk parastomataalne. Nad osalevad valverakkude liikumises. Moodustuvad valve- ja abirakud stomataalne aparaat.
Stoomide arv ja levik lehel või võrsel varieerub olenevalt taimeliigist ja elutingimustest. Nende arv ulatub tavaliselt mõnest kümnest kuni mitmesajani 1 ruutmm pinna kohta.
Kaitserakkude liikumismehhanism on väga keeruline ja eri liikide lõikes erinev. Enamikus taimedes, kui öösel ja mõnikord ka päeval on veevarustus ebapiisav, väheneb turgor kaitserakkudes ja vahe sulgub, vähendades seeläbi transpiratsiooni taset. Turgori suurenemisega stoomid avanevad. Arvatakse, et peamine roll nendes muutustes kuulub kaaliumiioonid. Kloroplastide olemasolu kaitserakkudes on turgori reguleerimisel hädavajalik. Kloroplastide esmane tärklis, muutudes suhkruks, suurendab rakumahla kontsentratsiooni. See soodustab vee sissevoolu naaberrakkudest ja kaitserakkude üleminekut elastsesse olekusse.
Stomataalsete avade kogupindala moodustab vaid 1-2% lehtede pindalast. Sellest hoolimata ulatub transpiratsioon avatud stomataalsete lõhedega 50-70% aurustumisest, mis on pindalalt võrdne avatud veepinnaga.