Як виглядає продиха. Характеристика основних екологічних груп рослин. Старіння листя та листопад
ТКАНИНИ. КЛАСИФІКАЦІЯ ТКАНИН.
В основі організації вищих рослин лежить принцип спеціалізації клітин, який полягає в тому, що кожна клітина організму виконує не всі властиві їй функції, а лише деякі, зате повніше і досконаліше.
Тканини- стійкі, закономірно повторювані комплекси клітин, подібні за походженням, будовою та пристосовані до виконання однієї або кількох функцій.
Існують різні класифікації тканин, але вони досить умовні.
Залежно від основної функції розрізняють кілька груп рослинних тканин.
1. Освітні тканиниабо меристеми,- мають здатність до поділу та формування всіх інших тканин.
2. Покривні тканини:
Первинні;
Вторинні;
Третинні.
3. Основні тканини- складають більшу частину тіла рослини. Розрізняють такі основні тканини:
асиміляційні (хлорофілоносні);
Запасні;
Повітроносні (аеренхіма);
Водоносні.
4. Механічні тканини(опорні, скелетні):
Колленхіма;
Склеренхіма.
5. Провідні тканини:
Ксилема (деревина) - тканина висхідного струму;
Флоема (луб) – тканина низхідного струму.
6. Видільні тканини:
Зовнішні:
Залізисті волоски;
Гідатоди - водяні продихи;
Нектарники;
Внутрішні:
Видільні клітини з ефірними оліями, смолами, дубильними речовинами;
Багатоклітинні вмістилища виділень, млечники.
За здатністю клітин до поділу розрізняють два типи тканин: освітні,або меристеми,і постійні- покривні, видільні, основні, механічні, провідні.
Тканину називають простий,якщо всі її клітини однакові за формою та функціями (паренхіма, склеренхіма, коленхіма). Складнітканини складаються з клітин, неоднакових за формою, будовою та функціями, але пов'язаних загальним походженням (наприклад, ксилема, флоема).
Існує також класифікація тканин, заснована на їхньому походженні (онтогенетична). Відповідно до цієї класифікації розрізняють первинні та вторинні тканини. З первинної меристеми, що знаходиться на верхівці втечі та кінчику кореня, а також із зародка насіння формуються первинні постійнітканини (епідерма, коленхіма, склеренхіма, асиміляційна тканина, епіблема). Клітини постійних тканин нездатні до подальшого поділу. З клітин спеціалізованої меристеми - прокамбія - формуються первинні провіднітканини (первинна ксилема, первинна флоема).
З вторинної меристеми - камбія - формуються вториннітканини: вторинна ксилема, вторинна флоема; з феллогену утворюються пробка, феллодерма, чечевички, що виникають при потовщенні стебла та кореня. Вторинні тканини, як правило, зустрічаються у голонасінних і дводольних покритонасінних рослин. Потужний розвиток вторинних тканин - деревини та лубу характерний для деревних рослин.
ОСВІТНІ ТКАНИНИ
Освітні тканинизавдяки постійному мітотичному поділу їх клітин забезпечують утворення всіх тканин рослини, тобто. фактично формують його тіло. Будь-яка клітина у своєму розвитку проходить три стадії: ембріональну, зростання та стадію диференціації (тобто набуття клітиною певної функції). У міру диференціації зародка первинна меристема зберігається тільки на верхівці майбутньої втечі (у конусі наростання) та на кінчику кореня – апікальні (верхівкові) меристеми.Зародок будь-якої рослини складається з клітин меристеми.
Цитологічна характеристика меристему.Типові ознаки найвиразніше виражені у верхівкових меристемах. Ці меристеми складені ізодіаметричними багатогранними клітинами, не розділеними міжклітинниками. Їхні оболонки тонкі, містять мало целюлози і здатні розтягуватися.
Порожнина кожної клітини заповнена густою цитоплазмою з відносно великим ядром, що займає центральне положення, і мітозом, що інтенсивно ділиться. У гіалоплазмі багато дифузно розкиданих рибосом, пропластид, мітохондрій та диктіосом. Вакуолей небагато, і вони дрібні. Провідні тканини утворюються з меристеми, що має прозенхімну форму і великі вакуолі - прокамбію і камбію. Клітини прокамбію в поперечному перерізі багатокутні, клітини камбію - прямокутні.
Клітини, що зберігають свої меристематичні властивості, продовжують ділитися, утворюючи нові і нові клітини, звані ініціалями.Частина дочірніх клітин диференціюється, перетворюючись на клітини різних тканин, їх називають похідними ініціалів.Клітини ініціалів можуть ділитися невизначено багато разів, а похідні ініціалів діляться один або кілька разів і перетворюються на постійні тканини.
За походженням розрізняють первинні та вторинні меристеми.
Первинні меристеми
Первинні меристеми походять безпосередньо з меристеми зародка і мають здатність до поділу. По положенню в рослині первинні меристеми можуть бути верхівковими (апікальними), вставковими (інтеркалярними) та бічними (латеральними).
Верхівкові (апікальні) меристеми- такі меристеми, які розташовуються у дорослих рослин на верхівках стебел та кінчиках коріння та забезпечують зростання тіла у довжину. Біля стебел у конусі наростання виділяють два меристематичні шари: туніку, з якої утворюються покривна тканина і периферична частина первинної кори, і корпус, з якого утворюються внутрішня частина первинної кори та центральний осьовий циліндр (рис. 2.3).
Мал. 2.3.Апікальні меристеми стебла: а- поздовжній зріз: 1 – конус наростання; 2 – зачаток листа; 3 - горбок пазушної нирки;
У кінчику кореня розрізняють три шари:
1) дерматоген, з якого утворюється первинна покривно-всмоктуюча тканина – ризодерма;
2) периблему, з якої розвиваються тканини первинної кори;
3) плером, що утворює тканини центрального осьового циліндра.
Бічні (латеральні) меристемиза походженням можуть бути первинними та вторинними, на поперечному зрізі осьових органів мають вигляд кілець. Прикладом первинної бічної меристеми є прокамбій і перицикл. З прокамбіяформуються камбій та первинні елементи судинно-волокнистих пучків (первинна флоема та первинна ксилема), при цьому клітини прокамбію безпосередньо диференціюються в клітини первинних провідних тканин.
Бічні меристеми розташовуються паралельно поверхні органу та забезпечують зростання осьових органів у товщину.
Вставкові (інтеркалярні) меристемичастіше первинні та зберігаються у вигляді окремих ділянок у зонах активного росту в різних частинах рослини (наприклад, в основі черешків листя, біля основ міжвузлів). В основі міжвузлів у злаків діяльність цієї меристеми веде до подовження міжвузлів, що забезпечує зростання стебла в довжину.
Вторинні меристеми
До вторинних відносять бічні та ранові меристеми.
Бічні (латеральні) меристемипредставлені камбіємі феллогеном.Вони формуються з промеристем (прокамбію) або постійних тканин шляхом їх дедиференціювання. Клітини камбію діляться перегородками, паралельними поверхні органу (периклінально). З клітин, відкладених камбієм назовні, розвиваються елементи вторинної флоеми, та якщо з клітин, відкладених всередину, - вторинної ксилемы. Камбій, що виник із постійних тканин шляхом дедиференціювання, називають додатковим.За будовою та функцією він не відрізняється від камбію, що виник з промеристем. Феллоген формується із постійних тканин, розташованих у субепідермальних шарах (під епідермою). Ділячись периклінально, феллоген відокремлює назовні майбутні клітини пробки (фелеми), а всередину - клітини фелодерми. Таким чином, феллоген формує вторинну покривну тканину – перидерму. Бічні меристеми розташовуються паралельно поверхні органу та забезпечують зростання осьових органів у товщину.
Раневі меристемиутворюються при пошкодженні тканин та органів. Навколо пошкодження живі клітини дедиференціюються, починають ділитися і цим перетворюються на вторинну меристему. Їхнє завдання - утворити щільну захисну тканину, що складається з паренхімних клітин, - каллюс.Ця тканина білуватого або жовтуватого кольору, її клітини мають великі ядра і досить товсті клітинні стінки. Каллюс виникає при щепленнях, забезпечуючи зрощення щепи з підщепою, і в основі черешків. У ньому можуть закладатися придаткові коріння та нирки, тому його використовують для одержання культури ізольованих тканин.
ПОКРОВНІ ТКАНИНИ
Первинна покривна тканина
До первинної покривноїтканини відносять епідерму власне епідермальних, навколоустьичних, замикаючих клітин продихом і трихом.
Пектинові речовини та целюлоза, що входять у клітинну стінку, можуть піддаватися ослизненню з утворенням слизіві камедей.Вони являють собою полімерні вуглеводи, споріднені з пектиновими речовинами, і відрізняються здатністю до сильного набухання при зіткненні з водою. Камеді в набряклому стані клейкі і можуть витягуватися в нитки, тоді як слизу сильно розпливаються і нитки не витягуються. Пектинові слизу зустрічаються у представників сімейств лілейних, хрестоцвітих, мальвових, липових, розоцвітих, на відміну від целюлозних слизів, що зустрічаються набагато рідше (наприклад, у орхідних).
Устячка
являють собою високоспеціалізовані утворення епідерми, що складаються з двох замикаючих клітин бобоподібної форми та устьичної щілини (своєрідного міжклітинника між ними). Є головним чином листі, але трапляються й у стеблі (рис. 2.6). 
Мал. 2.6.Будова продихи: а, б- Шкірка листа чебрецю (вид зверху і на поперечному зрізі); в- Шкірка зі стебла цереуса (родина кактусових); 1 – власне епідермальні клітини; 2 - замикаючі клітини продихи; 3 – устьична щілина; 4 - повітроносна порожнина; 5 – клітини хлорофілоносної паренхіми; А – кутикула; Б - кутикулярний шар - оболонка із суберином та воском; В – целюлозний шар стінки; Г - ядро з ядерцем; Д - хлоропласти
Стіни замикаючих клітин потовщені нерівномірно: стінки, спрямовані до щілини (черевні), значно потовщені порівняно зі стінками, спрямованими від щілини (спинними). Щілина може розширюватися і звужуватися, регулюючи транспірацію та газообмін. Під щілиною розташована велика дихальна порожнина (міжклітинник), оточена клітинами мезофілу листка.
Замикаючі клітини оточені навколоустьовими клітинами, які спільно утворюють устьичний комплекс(Рис. 2.7). Розрізняють такі основні типи устьичних комплексів: 
Мал. 2.7.Основні типи устьичного апарату: 1 – аномоцитний (у всіх вищих рослин, крім хвощів); 2 - діацитний (у папоротей та квіткових); 3 - парацитний (у папороті, хвощів, квіткових та гнетових); 4 – анізоцитний (тільки у квіткових); 5 - тетрацитний (переважно у однодольних); 6 - анциклоцитний (у папоротей, голонасінних та квіткових)
1) аномоцитний(безладний) - замикаючі клітини немає яскраво виражених околоустьичных клітин; характерний всім вищих рослин, крім хвойні;
2) анізоцитний(Нерівноклітинний) - замикаючі клітини продихи оточені трьома навколоустьичними клітинами, одна з яких набагато більша (або менше) інших;
3) парацитний(паралельно-клітинний) - одна навколоустьева клітина (або більше) розташована паралельно замикаючим;
4) діацитний(перехресно-клітинний) – дві навколоустьичні клітини розташовані перпендикулярно замикаючим;
5) тетрацитний(Від грец. tetra- чотири) - головним чином у однодольних;
Продихання розташовані на нижній стороні листа, але у водних рослин з плаваючим листям вони знаходяться тільки на верхній стороні листа. За формою клітин епідерми листа і розташування продихів можна відрізнити однодольне рослина від дводольного (рис. 2.8). Власне епідермальні клітини листка дводольних рослин в контурах хвилясті (рис. 2.9), а у однодольних вони витягнуті, ромбічної форми. 
Мал. 2.8.Розташування продихів на епідермі (вид з поверхні): а-дводольні рослини: 1 - Букви; 2 – кавун; б-однодольні рослини: 3 – кукурудза; 4 - ірис
Типи продихів за рівнем розташування щодо поверхні епідермісу можна підрозділити наступним чином.
1.7.1. Продихання, розташовані в одній площині з епідермісом. Найбільш поширений тип і зазвичай у описі мікроскопії лікарської рослинної сировини не вказується, тобто. цей пункт опускається. Діагностичними ознаками будуть або виступаючі, або занурені продихи.
1.7.2. Простирадла устячка - продихи, розташовані над епідермісом. Зазвичай при обертанні мікрогвинта мікроскопа (при опусканні об'єктиву) спочатку виявляються такі продихи, а вже потім з'являються клітини епідермісу, тому на фотографії відобразити їх з поверхні листа практично неможливо, так само як і зобразити на малюнку. В одній площині з епідермісом такі продихи можна побачити на поперечних зрізах, але для цього зріз повинен пройти через продихи, що важко отримати при їх рідкісному розташуванні на листі. Такі продихи характерні, наприклад, для листя мучниці.
1.7.3. Занурені продихи - продихи, занурені в епідерміс. При спостереженні під мікроскопом при обертанні мікрогвинта (при опусканні об'єктиву) спочатку чітко виявляються клітини епідермісу, потім стає можливим чіткіше побачити контури продихів. На фотографіях та малюнках препаратів із поверхні їх також важко відобразити. Зустрічаються в листі конвалії, листі вахти, листках евкаліпта. Іноді поглиблення, в яких розташовуються продихи, вистелені або прикриті волосками і називаються устьичними криптами.
1.8. Типи устьичних клітин
У літературі описується 19 типів, нами обрані лише ті, що використовуються в аналізі лікарської рослинної сировини.

Мал. 63.Типи устьичних клітин. А - сочевицеподібні; Б - сфероподібні; В - ковпачкоподібні; Г – човноподібні
1.8.1. Сочевицеподібні - 2 однакові клітини напівмісячної форми розташовані симетрично. На передній поверхні потовщення оболонки майже рівномірне. Щілина веретеноподібна (рис. 63, А). Тип устьичних клітин характерний більшості рослин.
1.8.2. Сфероподібні - дві однакові, сильно кругоподібно зігнуті клітини розташовані симетрично. На передній поверхні потовщення оболонки майже рівномірне. Щілина кругла (рис. 63, Б).
1.8.3. Ковпачкоподібні - дві однакові клітини напівмісячної форми у полярних частинах мають потовщення у вигляді ковпачка. Щілина веретеноподібна (рис. 63, В). Зустрічаються у наперстянок.
1.8.4. Човноподібні - Внутрішні стінки устьичних клітин потовщені. Щілина веретеноподібна (рис. 63, Г). Спостерігається у траві золототисячника, у листі вахти.
Механізм роботи продихів обумовлений осмотичними властивостями клітин. При освітленні поверхні листа сонцем у хлоропластах замикаючих клітин відбувається активний процес фотосинтезу. Насичення клітин продуктами фотосинтезу, цукрами тягне за собою активне надходження до клітин іонів калію, внаслідок чого концентрація клітинного соку в клітинах, що замикають, зростає. Виникає різниця концентрації клітинного соку навколоустьових та замикаючих клітин. З огляду на осмотичних властивостей клітин вода з околоустьичных клітин надходить у замикаючі, що веде до збільшення обсягу останніх і різкого зростання тургора. Потовщення «черевних» стінок клітин, що замикають, звернених до устьичної щілини, забезпечує нерівномірне розтягнення клітинної стінки; замикаючі клітини набувають вираженої бобоподібної форми, і устьична щілина відкривається. При зниженні інтенсивності фотосинтезу (наприклад, увечері) зменшується утворення цукрів у клітинах, що замикають. Приплив іонів калію припиняється. Концентрація клітинного соку в клітинах, що замикають, знижується в порівнянні з околоустьічними. Вода шляхом осмосу йде з клітин, що замикають, знижуючи їх тургор, в результаті вночі устьична щілина закривається.
Клітини епідерми щільно зімкнуті між собою, завдяки цьому епідерма виконує цілу низку функцій:
Перешкоджає проникненню хвороботворних організмів усередину рослини;
Захищає внутрішні тканини від механічних ушкоджень;
Регулює газообмін та транспірацію;
Через неї виділяються вода, солі;
Може функціонувати як всмоктувальна тканина;
бере участь у синтезі різних речовин, сприйнятті подразнень та у русі листя.
Трихоми - різні за формою, будовою та функціями вирости клітин епідерми: волоски, лусочки, щетинки тощо. Їх поділяють на криючі та залізисті. Залізисті трихоми,на відміну від криючих, мають клітини, що виділяють секрет. Кріючі волоски,утворюючи на рослині шерстистий, повстяний або інший покрив, відбивають частину сонячних променів і тим самим зменшують транспірацію. Іноді волоски знаходяться тільки там, де розташовані продихи, наприклад, на нижній стороні листа мати-й-мачухи. У деяких рослин живі волоски збільшують загальну поверхню, що випаровує, що сприяє прискоренню транспірації.
Розміри трихом значно варіюють. Найбільш довгі трихоми (до 5-6 см) покривають насіння бавовнику. Кроючі трихоми мають форму простих одно або багатоклітинних, розгалужених або зірчастих волосків. Трихоми, що кроять, можуть тривалий час залишатися живими або швидко відмирати, заповнюючись повітрям.
Від тріхом, що виникають лише за участю епідермальних клітин, відрізняються емергенці, в освіті яких беруть участь і глибше розташовані тканини субепідермальних шарів.
Анатомо-діагностичні ознаки, що мають найбільше значення та високу варіабельність при визначенні лікарської сировини. Волоски можуть бути прості та головчасті, які у свою чергу можуть бути одноклітинними та багатоклітинними. Багатоклітинні волоски можуть бути однорядними, дворядними та гіллястими.
ЛАБОРАТОРНА РОБОТА № 5
ВОДНИЙ ОБМІН. ЛИСТ ЯК ОРГАН ТРАНСПІРАЦІЇ
Мета роботи:вивчення найважливіших функціональних особливостей листка рослин як органу транспірації: будови та кількості продихів на листовій пластинці, механізму відкривання та закривання продихів, впливу різних речовин на рух продихів.
ТРАНСПІРАЦІЯ
Біологічне значення транспірації полягає, по-перше, у забезпеченні сталості внутрішньої температури аркуша. Це досягається поглинанням тепла водою при її випаровуванні листям. Енергія, необхідна для переведення молекули з рідкої фази в газоподібний стан без зміни температури, називається теплотою випаровування. Витрата тепла на випаровування води є засобом регуляції температури листя та попередження рослин від перегріву.
По-друге, транспірація, будучи верхнім кінцевим двигуном, забезпечує надходження води та елементів мінерального живлення до коріння. Встановлено наявність позитивної кореляції між інтенсивністю транспірації та надходження води та іонів. Якщо з рослини видалити листя, поглинання води корінням припиняється. У присмоктувальній дії транспіруючого листя можна переконатися, якщо помістити зрізану гілку в піпетку, заповнену водою, і опущену в чашку з ртуттю. Через деякий час можна спостерігати підняття ртуті в піпетці, що вказуватиме на значну силу, що присмоктує листя.
Таким чином, швидкість надходження води до коріння обумовлена інтенсивністю транспірації.
По-третє, транспірація запобігає виникненню надмірного тургорного тиску, що могло б призвести до руйнування клітин рослин.
По-четверте, процес транспірації перебуває у тісному зв'язку з фотосинтезом рослин, що було відзначено роботами К. А. Тимирязєва. Засвоєння СО 2 листям рослин відбувається через продихи, і воно залежить від ступеня насиченості листової тканини водою. Процес засвоєння води та вуглекислого газу є єдиним і нерозривним цілим.
Під інтенсивністю транспірації розуміють кількість води, що випарувалася за одиницю часу з одиниці листової поверхні. Зазвичай цей показник має розмірність – мг/дм 2 год. Кількість води, що випаровується рослинами, досить велика, і нерідко перевищує кількість опадів, що випали за вегетаційний період. Це перевищення компенсується осінньо-зимовими опадами. Так, наприклад, одна рослина соняшнику чи кукурудзи витрачає за літо 200-250 л води. Рослини пшениці на площі 1 га випаровують за літо близько 2 млн. літрів води, кукурудзи – понад 3 млн., а капусти – до 8 млн. л. У процесі утворення кілограма рослинної маси витрачається 300 л. Води.
Устьична транспірація регулюється ступенем відкритості продихів. Будова та розподіл їх залежить від видових та екологічних особливостей рослин. Продихання зустрічаються на всіх наземних частинах рослин, включаючи репродуктивні органи і навіть тичинкові нитки. Найбільш характерні продихи для листя. Найчастіше вони розташовуються на нижній стороні листя (у мезофітних рослин). Однак у ксерофітів вони трапляються і на верхній стороні листа.
Середня кількість продихів на 1 мм 2 площі коливається від 100 до 300. Розмір продихів не перевищує 20 мікрон в довжину і 8-15 мікрон завширшки. Загальна площа відкритих продихів складає 1% поверхні листа.
Встановлено, що дрібне верхівкове листя має більшу кількість продихів, ніж великі нижні. Частота продихів (число їх на одиницю площі) збільшується при переході від основи листа до його верхівки і від нижньої частини рослини до верхньої. У рослин посушливих місць проживання їх більше, але за розміром вони менші.
У більшості мезофітних рослин продихи розташовані на одному рівні з епідермальними клітинами, а у ксерофітних форм продихи розташовані нижче рівня епідермісу і називаються зануреними. У гігрофітів іноді замикаючі клітини розташовані вище за епідерміс. Такі продихи називаються піднятими.
Той чи інший тип будови продихів характерний для певних груп рослин, хоча в межах одного сімейства можуть іноді зустрічатися різні типи продихів. Незважаючи на значну площу, зайняту продихами, дифузія водяної пари через них становить 50-60% випаровування зі вільної поверхні. Встановлено, що швидкість дифузії через дрібні отвори пропорційна їхньому периметру, а не площі. Тому часткове змикання замикаючих клітин мало впливає на їх периметр, і рівень дифузії водяної пари через продихи не дуже різко падає.
Досвід 1. Спостереження за рухом продихів під мікроскопом.
Мета досвіду: визначити залежність роботи продихів від осмотично активних речовин.
Матеріали та обладнання: 5% розчин гліцерину, бритва, препарувальна голка, мікроскоп, скло предметне та покривне.
Рослини:листя (традесканції, тюльпана, гортензії або амариллісу, каланхое).
Газообмін між міжклітинниками листа та зовнішньою атмосферою регулюється продихами. Кожне продихи складається з двох замикаючих клітин, у яких стінки, що примикають до устьичної щілини, сильно потовщені, тоді як зовнішні частини оболонки залишаються тонкими. Неоднакова товщина зовнішніх і внутрішніх стінок призводить до того, що при зміні тургора клітини, що замикають, здатні кривитися або розпрямитися, відкриваючи або закриваючи при цьому устьичну щілину.
Хід роботи: виготовляють зрізи епідермісу листа вибраної рослини, які поміщають в 5% розчин гліцерину і витримують не менше 1 год. Роблять 10 промірів, знаходять середнє значення та обчислюють помилку середньої. Потім зрізи переносять із розчину гліцерину у воду і повторюють проміри устьичних щілин під мікроскопом. Результати заносять до таблиці 1.
Таблиця 1
Ступінь розкриття устьичної щілини в різних середовищах
|
Рослина, орган |
№ проміру |
Ступінь відкриття устьичної щілини |
|
|
Гліцерин | |||
|
Лист рослини | |||
Завдання:зробити висновок про вплив гліцерину та води на відкриття та закриття продихів.
Досвід 2. Визначення стану продихів і міжклітинників методом Моліша
Мета досвіду: визначить вплив зовнішніх умов на стан продихів та інтенсивність транспірації.
Матеріали та обладнання: ксилол (у крапельниці), етиловий спирт (у крапельниці); бензол (у крапельниці), піпетки.
Рослина: свіже або підв'яле листя рослин, листя рослин, що знаходилися в темряві.
Міжклітини листа зазвичай бувають заповнені повітрям, завдяки чому при розгляді світ лист видається матовим. Якщо зробити інфільтрацію, тобто. Заповнення міжклітинників будь-якою рідиною, то відповідні ділянки листа стають прозорими.
Визначення стану продихів методом інфільтрації засноване на здатності рідин, що змочують клітинні оболонки, проникати в силу капілярності через відкриті устьичні щілини в найближчі міжклітини, витісняючи з них повітря, в чому легко переконатися по появі на листі прозорих плям. Різні рідини здатні проникати в устьичні щілини, відкриті різною мірою: ксилол легко проникає через слабко відкриті продихи, бензол - через продихи відкриті середньо, а етиловий спирт здатний проникати тільки через широко відкриті продихи.
Даний метод, запропонований Молішом, дуже простий і цілком застосовний для роботи в польових умовах.
Хід роботи. На нижню поверхню листа нанести окремо маленькі краплі бензолу, ксилолу та етилового спирту. Тримати лист у горизонтальному положенні до повного зникнення крапель, які можуть або випаруватися, або проникнути всередину листа, та розглянути лист на світло.
Дослідити листя, витримані в різних умовах (свіжі та підв'ялі, освітлені та затінені тощо). Щоразу досліджувати 2-3 аркуші.
Таблиця 2
Вплив зовнішніх умов на ступінь відкриття продихів
Завдання:Результати записати в таблицю 2, відзначаючи ступінь відкритості продихів: широко, середньо, слабо. Зробити висновок про вплив зовнішніх умов на устьичні рухи.
Досвід 3. Визначення стану продихів за допомогою відбитків по Молотковському.
Мета роботи:визначення роботи продихів в залежності від освітленості.
Матеріали та обладнанняКабіна: безбарвний лак для нігтів, тонка скляна паличка, пінцет, мікроскоп, окуляр-мікрометр, об'єкт-мікрометр.
Рослини:кімнатні рослини, листя яких за 2-3 години до заняття закривають світлонепроникним чохлом.
На поверхню листа наносять тонкий мазок лаку. Після випаровування розчинника утворюється плівка, на якій друкується епідерміс з продихами. Розглядаючи отримані відбитки в мікроскоп, можна визначити кількість та розмір продихів, виміряти ширину устьичних щілин. Даний метод можна використовувати не тільки для лабораторних, але і для польових досліджень (в останньому випадку слід зберігати відбитки до визначення в пробірках з водою). Для дослідження листя, продихи яких розташовані в поглибленнях епідермісу (наприклад, у олеандра), цей метод не застосовується, т.к. у такого листя відбитки не виходять.
Хід роботи. Нанести на нижню сторону листа за допомогою скляної палички краплю розчину лаку та швидко розмазати тонким шаром. Після висихання зняти плівку пінцетом, помістити на предметне скло та розглянути при великому збільшенні. Вставити в мікроскоп окулярний мікрометр і виміряти ширину і довжину устьичної щілини не менше, ніж у 10 продихів і обчислити середні величини.
Визначити ціну поділу окулярного мікрометра. Для цього помістити на предметний столик мікроскопа об'єкт-мікрометр, кожен розподіл якого дорівнює 0,01 мм або 10 мкм. Повертаючи окуляр поєднати обидві шкали так, щоб їх шкали були паралельні та одна перекривала іншу. Визначення ціни розподілу окулярного мікрометра проводиться у принципі ноніуса, тобто. поєднують одну з рис шкали окулярного і об'єктивного мікрометра і знаходять наступне поєднання. Знайти збігаються лінії і визначити, скільки поділів окулярного мікрометра відповідають поділам об'єкт-мікрометра, що знаходиться між сполученими точками. Ціна поділу окулярного мікрометра визначається за формулою:
Ціна поділу = · 10 мкм/А.
Помноживши довжину і ширину продихів отворів, виражених у поділах окулярного мікрометра, на ціну одного поділу, знайти абсолютні розміри продихових щілин. Обчислити площу устьичної щілини з деяким наближенням шляхом множення довжини на ширину.
Дослідити листя різних ярусів однієї і тієї ж рослини, а також добре освітлені та затінені. Результати записати до таблиці 3.
Таблиця 3
Вплив освітленості на розміри устьичних отворів
Завдання:зробити висновки про вплив ярусності та умов висвітлення на розміри устьичних отворів.
Хоча вчені давно знали про випаровування води поверхнею листа, першим, хто спостерігав устячка, був італійський натураліст Марчелло Мальпігі, який це відкриття опублікував у 1675 році у своїй роботі Anatome plantarum. Однак він не зрозумів їхню справжню функцію. У той же час його сучасник Неемія Грю розвинув гіпотезу про участь продихів у вентиляції внутрішнього середовища рослини і порівняв їх з трахеями комах. Прогрес у вивченні настав у XIX столітті, і тоді ж, у 1827 році, швейцарським ботаніком Декандолем було вперше використано слово „stoma“. Вивченням продихів у той час займалися Гуго фон Моль, який відкрив основний принцип відкривання продихів і Симон Швенденер, що класифікував продихи за типом їх конструкції.
Деякі аспекти функціонування продихів продовжують інтенсивно вивчатися і в даний час; матеріалом в основному служать Коммеліна звичайна ( Commelina communis), Боб садовий ( Vicia faba), Кукурудза цукрова ( Zea mays) .
Будова
Розміри продихи (довжина) коливаються в межах 0,01-0,06 мм (більше бувають продихи поліплоїдних рослин і у листя, що росте в тіні. Найбільші продихи були виявлені у вимерлої рослини Zosterophyllum, 0,12 мм (120 мкм). Час складається з пари спеціалізованих клітин, званих замикаючими ( cellulae claudentes), які регулюють ступінь відкритості пори, між ними розташовується устьична щілина ( porus stomatalis). Стіни замикаючих клітин потовщені нерівномірно: спрямовані до щілини (черевні) товщі стінок, спрямованих від щілини (спинних). Щілина може розширюватися і звужуватися, регулюючи транспірацію та газообмін. Коли води мало, клітини, що замикають, щільно прилягають одна до одної і устьична щілина закрита. Коли води в клітинах, що замикають багато, то вона тисне на стінки і більш тонкі стінки розтягуються сильніше, а товстіші втягуються всередину, між замикаючими клітинами з'являється щілина. Під щілиною розташована подустьична (повітряна) порожнина, оточена клітинами м'якоті листа, через яку безпосередньо відбувається газообмін. Повітря, що містить діоксид вуглецю (вуглекислий газ) і кисень, проникає всередину тканини листа через ці пори, і далі використовується в процесі фотосинтезу та дихання. Надлишковий кисень, вироблений у процесі фотосинтезу внутрішніми клітинами листка, виходить назад у довкілля через ці пори. Також у процесі випаровування через пори виділяються пари води. Клітини епідермісу, що примикають до замикаючих, отримали назву супроводжуючих (побічних, сусідніх, навколоустицьких). Вони беруть участь у русі замикаючих клітин. Клітки, що замикають і супроводжують, утворюють устьичний комплекс (устьичний апарат). Наявність або відсутність продихів (видимі частини продихів називають устьичними лініями) часто використовують при класифікації рослин.
Типи продихів
Число супроводжуючих клітин та їх розташування щодо устьичної щілини дозволяють виділити ряд типів продихів:
- аномоцитно-супровідні клітини не відрізняються від інших клітин епідермісу, тип дуже звичайний для всіх груп вищих рослин, за винятком хвойних;
- діацитний - характеризується тільки двома клітинами, що супроводжують, загальна стінка яких знаходиться під прямим кутом до замикаючих клітин;
- парацитний - супроводжуючі клітини розташовуються паралельно замикаючої та устьичної щілини;
- анізоцитний - замикаючі клітини оточені трьома супроводжуючими, одна з яких помітно більша або дрібніша за інші, такий тип виявлений тільки у квіткових рослин;
- тетрацитний - чотири супроводжуючі клітини, характерний для однодольних;
- енциклоцитний - супровідні клітини утворюють вузьке колесо навколо замикаючих клітин;
- актиноцитний - кілька клітин, що супроводжують, радіально розходяться від замикаючих клітин;
- перицитний - замикаючі клітини оточені однією побічною супроводжуючою клітиною, продихом не з'єднане з супроводжуючою клітиною антиклінальною клітинною стінкою;
- десмоцитний - замикаючі клітини оточені однією супроводжуючою клітиною, продихом з'єднано з нею антиклінальною клітинною стінкою;
- полоцитний – замикаючі клітини оточені однією супроводжуючою не повністю: до одного з устьичних полюсів примикає одна або дві епідермальні клітини; продиху прикріплено до дистальної сторони єдиної клітини, що супроводжує, має U-подібну або підковоподібну форму;
- стефаноцитний - продих, оточений чотирма або більше (зазвичай п'ять-сім) слабодиференційованими супроводжуючими клітинами, що утворюють більш-менш виразну розетку;
- латероцитний – такий тип устьичного апарату розглядається більшістю ботаніків як проста модифікація аномоцитного типу.
Розташування продихів
Дводольні рослини, як правило, у нижній частині листа мають більше продихів, ніж у верхній. Це пояснюється тим, що верхня частина горизонтально-розташованого листа, як правило, краще освітлена, і менша кількість продихів у ній перешкоджає надмірному випаровування води. Листя з продихами, розташованими на нижній стороні, називаються гіпостоматичними.
У однодольних рослин наявність продихів у верхній і нижній частині листа по-різному. Дуже часто листя однодольних рослин розташоване вертикально, і в цьому випадку кількість продихів на обох частинах листа може бути однаково. Таке листя називається амфістоматічним.
У плаваючого листя на нижній частині листа продихів відсутні, так як вони можуть вбирати воду через кутикулу . Листя з продихами, розташованими на верхній стороні, називаються епістоматичними. У підводного листя продихів відсутні зовсім.
Продихання хвойних рослин зазвичай заховані глибоко під ендодермою, що дозволяє сильно знизити витрату води взимку на випаровування, а влітку - під час посухи.
У мохів (виключення антоцеротові) справжні продихи відсутні.
Продихи також відрізняються за рівнем розташування щодо поверхні епідермісу. Деякі з них розташовані нарівні з іншими епідермальними клітинами, інші підняті вище або занурені нижче за поверхню. У однодольних, листя яких росте переважно в довжину, продихи утворюють правильні паралельні ряди, тоді як у дводольних вони розташовуються безладно.
Вуглекислий газ
Так як вуглекислий газ є одним із ключових реагентів у процесі фотосинтезу, у більшості рослин продихи у денний час відкриті. Проблема полягає в тому, що при вході повітря поєднується з парами води, що випаровуються з листа, і тому рослина не може отримати вуглекислий газ, одночасно не втративши деяку кількість води. У багатьох рослин існує захист від випаровування води у вигляді закупорюючих продихів воскових відкладень.
Особливого значення в житті рослини мають продихи, що належать до системи епідермальної тканини. Будова продихів настільки своєрідна і значення їх настільки велике, що їх слід розглянути особливо.
Фізіологічне значення епідермальної тканини має подвійний, значною мірою суперечливий характер. З одного боку, епідерміс структурно пристосований до захисту рослини від висихання, чому сприяє щільне змикання епідермальних клітин, утворення кутикули та відносно довгих волосків, що криють. Але з іншого боку, епідерміс повинен пропустити крізь себе маси пари води і різних газів, що спрямовуються у взаємно протилежних напрямках. Газо- та парообмін за деяких обставин можуть бути дуже інтенсивними. У рослинному організмі зазначене протиріччя успішно дозволяється з допомогою продихів. Устячка складається з двох своєрідно змінених епідермальних клітин, з'єднаних між собою протилежними (за своєю довжиною) кінцями і званих замикаючими клітинами. Міжклітинник між ними зветься устьичній щілини.
Клітки, що замикають, називаються так тому, що вони шляхом активної періодичної зміни тургору змінюють свою форму таким чином, що устьична щілина то відкривається, то закривається. Для цих устьичних рухів велике значення мають дві особливості. По-перше, клітини, що замикають, на відміну від інших клітин епідермісу містять хлоропласти, в яких на світлі відбувається фотосинтез і утворюється цукор. Накопичення цукру як осмотично діяльної речовини викликає зміну тургорного тиску клітин, що замикають, в порівнянні з іншими клітинами епідермісу. По-друге, оболонки замикаючих клітин потовщуються нерівномірно, тому зміна тургорного тиску викликає нерівномірну зміну об'єму цих клітин, отже, зміна їх форми. Зміна форми замикаючих клітин і викликає зміну ширини устьичной щілини. Пояснимо це на прикладі. На малюнку зображено один із типів продихів дводольних рослин. Саму зовнішню частину продиху складають плівчасті виступи, утворені кутикулою, іноді нікчемні, а іноді досить значні. Вони обмежують від зовнішньої поверхні невеликий простір, нижню межу якого становить сама щілина продихи, що носить назву переднього дворика продихи. За щілиною продиху, ковнутрі, розташовано ще один невеликий простір, відмежований невеликими внутрішніми виступами бічних стінок замикаючих клітин, зване внутрішнім двориком продихом. Внутрішній дворик безпосередньо відкривається у великий міжклітинник, званий повітряною порожниною.
На світлі в клітинах, що замикають, утворюється цукор, він відтягує воду від сусідніх клітин, тургор замикаючих клітин збільшується, тонкі місця їх оболонки розтягуються сильніше, ніж товсті. Тому опуклі виступи, що випинаються в щілину продиху, стають плоскими і продихом розкривається. Біли цукор, наприклад, уночі переходить у крохмаль, то тургор у замикаючих клітинах падає, це викликає ослаблення розтягнутості тонких ділянок оболонки, вони випинаються назустріч один одному і продиху закривається. У різних рослин механізм закривання та відкривання щілини продихів може бути різним. Наприклад, у злаків та осок замикаючі клітини мають розширені кінці та звужені в середній частині. Оболонки у середніх частинах клітин потовщені, тоді як розширені кінці зберігають тонкі целюлозні оболонки. Збільшення тургору викликає набухання кінців клітин і внаслідок цього відходження друг від друга прямих медіанних елементів. Це і призводить до розкриття продиху.
Особливості механізму роботи устьичного апарату створюються як формою і будовою замикаючих клітин, і участю у ньому сусідніх з продихами клітин епідермісу. Якщо клітини, що безпосередньо примикають до продихів, відрізняються за своїм виглядом від інших клітин епідермісу, їх називають супроводжуючими клітинами продихів.
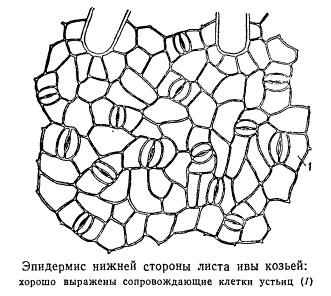
Найчастіше супроводжуючі та замикаючі клітини мають загальне походження.
Замикаючі клітини продихи або трохи підняті над поверхнею епідермісу, або, навпаки, опущені в більш-менш глибокі ямки. Залежно від положення замикаючих клітин по відношенню до загального рівня поверхні епідермісу дещо змінюється і механізм регулювання ширини устьичной щілини. Іноді замикаючі клітини продиху одревесневают і тоді регулювання відкривання устьичної щілини визначається діяльністю сусідніх епідермальних клітин. Розширюючись і зіщулюючись, тобто змінюючи свій обсяг, вони захоплюють замикаючі клітини, що примикають до них. Однак нерідко продихання з одревесневшими замикаючими клітинами зовсім не закриваються. У таких випадках регуляція інтенсивності газо- і парообміну здійснюється інакше (шляхом так званого підсихання, що починається). У продихах з одревесневшими замикаючими клітинами кутикула часто покриває досить потужним шаром не тільки всю устьичну щілину, але поширюється навіть на повітряну порожнину, вистилаючи її дно.
У більшості рослин продихи є на обох сторонах листа або тільки на нижній стороні. Але є й такі рослини, у яких продихи утворюються лише на верхній стороні листка (на листі, що плаває на поверхні води). Як правило, продихів на листі більше, ніж на зелених стеблах.
Число продихів на листі різних рослин дуже різне. Наприклад, число продихів на нижній стороні листа багаття безостого дорівнює в середньому 30 на 1 мм 2 , у соняшнику, що росте в тих же умовах, - близько 250. У деяких рослин налічується до 1300 продихів на 1 мм 2 .
У екземплярів одного й того ж виду рослин густота та розміри продихів сильною мірою залежать від екологічних умов. Наприклад, на листі соняшника, вирощеного на повному світлі, на 1 мм 2 листової поверхні в середньому припадало 220 продихів, а у екземпляра, вирощеного поряд з першим, але при невеликому затіненні - близько 140. На одній рослині, що вирощується на повному світлі, густота продихів зростає від нижнього листя до верхнього.
Число і розміри продихів сильно залежать не тільки від умов росту рослини, але і від внутрішніх взаємовідносин життєвих процесів у самій рослині. Ці величини (коефіцієнти) є найчутливішими реагентами на кожну комбінацію факторів, що зумовлюють зростання рослини. Тому визначення густоти та розмірів продихів листя рослин, вирощених у різних умовах, дає деяке уявлення про характер взаємовідносини кожної рослини з навколишнім середовищем. Усі методи визначення розмірів та кількості анатомічних елементів у того чи іншого органу відносяться до категорії кількісно-анатомічних методів, якими іноді користуються при екологічних дослідженнях, а також для характеристики сортів культурних рослин, тому що кожному сорту будь-якої культурної рослини властиві певні межі розмірів та кількість анатомічних елементів на одиницю площі. Методи кількісної анатомії можуть бути застосовані з великою користю як у рослинництві, так і в екології.
Поряд з продихами, призначеними для газо-і парообміну, існують ще продихи, через які виділяється вода не у вигляді пари, а в краплинно-рідкому стані. Іноді такі продихи цілком подібні до звичайних, тільки трохи більші за них, а замикаючі клітини їх позбавлені рухливості. Дуже часто у такого продиху в цілком зрілому стані замикаючі клітини відсутні і залишається лише отвір, що виводить воду назовні. Продихання, що виділяють крапельно-рідку воду, називають водними, а всі утворення, що беруть участь у виділенні крапельно-рідкої води, - гідатодами.
Будова гідатод різноманітна. Одні гідатоди мають під отвором, що виводить воду, паренхіму, що бере участь у передачі води від водопровідної системи та у виділенні її з органу; в інших гідатод водопровідна система безпосередньо підходить до вивідного отвору. Особливо часто гідатоди утворюються на першому листі проростків різних рослин. Так, у вологу та теплу погоду молоде листя злаків, гороху та багатьох лугових трав крапля за краплею виділяють воду. Це явище можна спостерігати в першу половину літа рано вранці кожного погожого дня.
Найбільш добре виражені гідатоди розташовані по краях листя. Нерідко одну або кілька гідатод несе кожен із зубчиків, що обертають краї листя.
Для зіткнення аркуша з атмосферою є пори - продихи. Устьиці - це отвір (щілина), обмежена двома замикаючими клітинами. Продихи зустрічаються у всіх наземних органів рослини, але найбільше у листя. Кожна замикаюча клітина продихи на відміну від клітин епідермісу має хлоропласту. У них відбувається фотосинтез,хоча з меншою інтенсивністю, ніж у клітинах мезофілу. Продихання - одне з оригінальних пристосувань, що володіють здатністю відкриватися і закриватися в залежності від насиченості замикаючих клітин водою. Зазвичай устьичні отвори обмежені двома замикаючими клітинами, стінки яких нерівномірно потовщені. У дводольних рослин замикаючі клітини бобовидної, або півмісячної форми, при цьому їх внутрішні прилеглі один до одного клітинні стінки більш товсті, а зовнішні - більш тонкі. Протопласти замикаючих клітин пов'язані в єдине ціле перфораціямив основі загальних стінок, що межують. Коли води мало, клітини, що замикають, щільно прилягають одна до одної і устьична щілина закрита. Коли води в клітинах, що замикають, багато, то вона тисне на клітинні стінки, і більш тонкі стінки розтягуються сильніше, а товстіші втягуються всередину, між замикаючими клітинами з'являється щілина.
Останнім часом доведено, що для руху продихів велике значення має також розташування мікрофібрил целюлози. Якщо зазвичай в клітинах листя целюлозні фібрили орієнтовані в довжину і в цьому напрямі потовщені, то в клітинах, що замикають, продихів мікрофібрили організовані радіально, що посилює стійкість до процесу розтягування.
У злаківбудова замикаючих клітин дещо інше. Вони представлені двома подовженими клітинами, на кінцях яких більш тонкі стінки. При насиченні водою тонші стінки на кінцях розтягуються і розсувають замикаючі клітини, завдяки чому утворюється щілина.
Число устьичних отворів коливається в залежності від виду рослин від 10 до 600 на 1 мм2 листа. У багатьох рослин (75% видів), у тому числі для більшості деревних, продихи розташовані на нижній стороні листа. Діаметр устьичних щілин становить лише 3-12 мкм. Продихання з'єднують внутрішні простори листа із зовнішнім середовищем. Вода надходить у лист через мережу жилок, у яких розташовані судинні елементи. Можливі три шляхи випаровування:
- через устячка - устьична,
- кутикулу - кутикулярна,
- через чечевички – лентикулярна транспірація.
Вперше розмежування на кутикулярну та устьичну транспірацію було введено у 1877 році.
Основні типи устьичного апарату листя рослин.
- аномоцитний (у всіх вищих рослин, крім хвощів),
- діацитний (у папороті квіткових),
- парацитний (у папоротей, хвощів, квіткових та гнетових),
- анізоцитний (тільки у квіткових),
- тетрацитний (головним чином у однодольних),
- енциклоцитний (у папоротей, голонасінних і квіткових).
Продихання є високоспеціалізовані утворення епідерми, що складаються з двох замикаючих клітин, між якими є своєрідний міжклітинник, або устьична щілина.
Щілина може розширюватися і звужуватися, регулюючи транспірацію та газообмін. Під щілиною розташовується дихальна, або повітряна порожнина, оточена клітинами м'якоті листка. Клітини епідерми, що примикають до замикаючих, дістали назву побічних, або навколоустицьких. Вони беруть участь у русі замикаючих клітин. Замикаючі та побічні клітини утворюють устьичний апарат.
Число і розподіл продихів на листі або втечі варіюють залежно від виду рослин та умов життя. Число їх зазвичай коливається від кількох десятків до кількох сотень на 1 кв.мм поверхні.
Механізм руху замикаючих клітин дуже складний і неоднаковий у різних видів. У більшості рослин при недостатньому водопостачанні в нічний годинник, а іноді і вдень тургор в клітинах, що замикають, знижується і щілина замикається, знижуючи тим самим рівень транспірації. З підвищенням тургору продихи відкриваються. Вважають, що головна роль цих змін належить іонам калію.Істотне значення у регуляції тургору має присутність у замикаючих клітинах хлоропластів. Первинний крохмаль хлоропластів, перетворюючись на цукор, підвищує концентрацію клітинного соку. Це сприяє припливу води із сусідніх клітин та переходу замикаючих клітин у пружний стан.
Загальна площа продихових отворів становить лише 1-2% площі листа. Незважаючи на це, транспірація при відкритих устьичних щілинах досягає 50-70% випаровування, що дорівнює площі відкритої водної поверхні.


















