Kloroplastid on rohelised plastiidid. Millistes plastiidides on pigment klorofüll. Division Sinivetikad. Organisatsiooni tunnused, eluvormid, pigmendid, elutsükkel
PLASTIIDID
Plastiide leidub alati protoplasmas, on füüsikaliste ja keemiliste omaduste poolest sellele lähedased, tekivad ainult plastiididest. Nad on võimelised kasvama ja paljunema jagunemise teel, nad võivad moodustada teatud pigmente oma kehas (stroomas) ja moodustada tärklist stroomas. Sõltuvalt teatud pigmentide sisaldusest leitakse kõrgemate taimede põhiplastiidide värvust ja funktsioone: a) rohelised plastiidid (kloroplastid), b) punased ja kollased (kromoplastid) ja c) värvitud (leukoplastid).
Kõik plastiidid on ilmselt sarnase struktuuriga; paremini uurinud kloroplastide struktuuri.
1 - niidipuur Zygnema cruciatum; 2 - eraldi rakk spirogyra niidist; P- pürenoidid.
X- kromatofoor; P- tärklisega pürenoidid; I- tuum.
Kloroplastid. Kõrgemate taimede kloroplastid (nimetatakse ka klorofülli teradeks) on kuju poolest mõnevõrra sarnased läätsedega: plaani järgi on kloroplastil kuju, mis on enam-vähem ringilähedane, profiilis vaadatuna meenutab see ellipsi. Kui klorofülli terad asuvad tihedas kihis, siis üksteisele vajutades omandavad nad nurga kuju. Klorofülli terade arv erinevates rakkudes on väga varieeruv. Näiteks kastooroa lehtede rakkudes on klorofülli terade arv 10 kuni 36, rakkudes Elodea densa- 26 kuni 32. Klorofülli terade läbimõõt on 4-9 μ.
Suuruse poolest on klorofülliterad vähem mitmekesised kui tuumad ja veelgi enam rakud ise, kuigi mõned autorid märgivad, et suurtes rakkudes on klorofülliterad suuremad. Kui I. I. Gerasimov sai spirogyra kultuurides suuri rakke kahekordse tuuma massiga, siis sellistes rakkudes olid ka spiraalsed kloroplastid suuremad kui tavalistes ja nende arv kasvas 8-lt 12-13-le (joonis 18). Erakordselt suuri kloroplaste on täheldatud Peperomia metallica: läbimõõt
kloroplastid ulatuvad 24 μ-ni, kuid siin tuleb märkida, et nende arv rakus on väga väike - neid on ainult 4.
Kloroplastid võivad muuta kuju ja suurust. Mõned muudatused sõltuvad pindpinevusest; selle suurenedes pinna suurus väheneb ja plastiidi kuju läheneb sfäärilisele - plastiid on "ümardatud"; pindpinevuse vähenemisega plastiid pikeneb. Kloroplastide kuju võib sõltuvalt valgustusest muutuda; näiteks varjutatud riitsinusoa lehtedes muutuvad kloroplastid peaaegu isodiameetriliseks (maksimaalne suurus ~6,3 µ ja minimaalne suurus ~5,7 µ); eredas valguses muudavad nad kuju läätsekujuliseks (diameetriga ~8,3 μ ja paksusega ~3,6 μ).
Vesikatku (elodea) kultuuris osutusid erinevatel temperatuuridel kõrgemal temperatuuril kasvanud lehtede klorofülliterad peaaegu poole suuremaks.
Vetikate kloroplastid – kromatofoorid – on väga mitmekesise kujuga (joon. 28, 29).
Paljude vetikate (joonis 28, 29) ja mõnede perekonda kuuluvate maksarohtude värvilistes plastiidides - kromatofoorides Anthoceros, on spetsiaalsed, viskoossed-vedelad kehad, kõige sagedamini ümarad või nurgelised; need väikesed kehad, mida nimetatakse pürenoidideks, on rikkad valguainete poolest, kuid ei sisalda nukleiine. Väikesed tärkliseterad paiknevad tavaliselt pürenoidide ümber; need tärklise terad tekivad rakus esimestena ja kaovad viimasena. Pürenoidid tekivad olemasolevate jagunemisel, kuid võivad tekkida ka rakus uuesti.
Kloroplastid sisaldavad stroomas nelja pigmenti: kahte rohelist (klorofüll a ja klorofüll b), oranžikaspunane (karoteen või muidu karoteen) ja kollane (ksantofüll).
Keemilise koostise järgi on klorofüll dikarboksüülhappe klorofülliini ja kahe alkoholi - metüüli ja fütooli ester.
Klorofüll a erineb klorofüllist b vesiniku ja hapniku aatomite arvu järgi.
Klorofüll a on sinaka varjundiga, klorofüll b- kollakas. Klorofülli molekulmass on ~900.
IP Borodin, töödeldes taime roheliste osade sektsioone slaidil etüülalkoholiga, saadi pärast preparaadi aeglast kuivatamist tumerohelised või peaaegu mustad kristallid kolme- või kuusnurksete plaatide ja tetraeedrite kujul. Hiljem selgus, et tegemist on klorofülli kristallidega, mille molekulides on fütoolirühm asendatud etüüliga.
Keskne koht klorofülli molekulides a ja b hõivab magneesiumiaatomi, mis on seotud 4 lämmastikuaatomiga.
Üks klorofülli tera sisaldab 6% klorofülli; ülejäänu on vesi, valgud, lipiidid jne.
Klorofüllidega kaasnevatest pigmentidest on oranžikaspunane karoteen küllastumata süsivesinik valemiga C 40 H 56 ja kollane ksantofüll (C 40 H 56 O 2) on kahehüdroksüülne alkohol, justkui karoteeni oksüdatsiooni produkt. Karoteen ja ksantofüll kuuluvad suurde karotenoidide rühma – kollased, oranžid ja punased pigmendid, mille mitmeid teisi esindajaid leidub ka taimedes. Kloroplasti pigmentide kogumit nimetatakse mõnikord "klorofülliks" selle sõna laiemas tähenduses.
Roheliste lehtede ekstrakt ja vähemal määral ka lehed ise näitavad fluorestsentsi nähtust. Läbiva valguse korral paistab kapuuts roheline, langevas valguses aga kirsipunane. Väga harvade eranditega ei teki katteseemnetaimede elundites valguse puudumisel kloroplastides rohelisi pigmente või neid tekib ebaolulises koguses; valguses muutuvad nad kiiresti roheliseks.
Kloroplastides toimub kompleksne fotosünteesi protsess - päikesevalguse energia mõjul süsihappegaasist ja veest süsivesikute moodustumine.
Fotosünteesi protsessi lõpptulemust võib kujutada järgmise reaktsioonina:
6CO2 + 6H2O + 674 kcal→ C6H12O6 + 6O2
Saadud süsivesik polümeriseeritakse tavaliselt tärkliseks vastavalt skeemile:
n C6H12O6 → (C6H10O5) n + n H2O

Riis. 30. Samblalehe klorofülliterad funaria hydrometrica:
1 - mitme rakuga täiskasvanud lehe ala (plaanis); protoplasma seinakihis on klorofülliterad väikeste tärkliseteradega (märgitud valgega); 2-10 - üksikud tärklisega klorofülliterad: 2 - noor 3 - küpsem 9 ja 10 - jagamine, 4, 5 ja 6 - tärklisega täidetud, 7 - noored, vees paistes, 8 - vees hägune ja maha jäänud tärkliseterad.
Tärklis ladestub kloroplastidesse väikeste teradena assimilatsioon, ehk autohtoonne, tärklis (joon. 30, 32).
Mõnes taimes, enamasti üheidulehelistes, assimilatsioonitärklist tavaliselt ei moodustu (v.a stoomi kaitserakkudes) ja fotosünteesi saadus on glükoos. Kui suhkrut moodustavate taimede (suhkruroog, suhkrupeet) kloroplastides on atmosfääris oluliselt suurenenud CO 2 sisaldus, ilmub valgusesse tärklis.
Plastiidide kui bioloogiliselt aktiivsete süsteemide fotosünteesivõime ja mitmed muud omadused on seletatavad ensüümide olemasoluga kloroplastides. See ülimalt keeruline ensümaatiline süsteem pakub mitte ainult

kogu fotosünteesi protsessi, aga ka assimilatsiooniproduktide väljavoolu kloroplastist. Selle süsteemi koostis sisaldab plastiidide rohelist pigmenti - klorofülli.
Kloroplasti siseehituse üle oli palju vaidlusi, esitati palju teooriaid, kuid ainult elektronmikroskoobi kasutamine võimaldas selle submikroskoopilist ehitust täpsemalt uurida (joon. 31, 32). Praegu arvatakse, et kõrgemate taimede kloroplastid on lamellstruktuuriga. Stroomaplaadid vahelduvad plaatidega, mis koosnevad klorofülli sisaldavatest graanulitest (graanulitest). Graanulite ja stroma vaheline seos ei ole praegu täiesti selge.
Küsimuste selgitamisel klorofülli rolli kohta fotosünteesi protsessis ja päikesespektri erinevate piirkondade kiirte tähtsuse kohta selles protsessis kuulub K. A. Timirjazevi suur teene. Timirjazev uuris klorofülli kui "ühendust päikese ja elu vahel" ja klorofülli tera kui seda fookust, seda punkti kosmoses, kus keemiliseks energiaks muutuv päikesekiir muutub kogu elu allikaks Maal.
Timiryazev leidis, et punased kiired neelduvad kõige intensiivsemalt (lainepikkusega alates

Paremal - moodustunud kloroplast, mille stroomas on näha kolm läätsekujulist tärklisetera.
730 kuni 680 mμ) ja vähemal määral spektri sinakasvioletse osa kiired (lainepikkusega 470 mμ ja vähem).
Sügisel, enne lehtede langemist, muutuvad klorofülliterad kollaseks; see kollasus sõltub sellest, et rohelised pigmendid hävivad enne kollaseid; kollasus on seotud ainete väljavooluga lehelabast aksiaalorganitesse.
Kromoplastid. Kromoplastid on plastiidid, mis sisaldavad karotenoidpigmente (karoteeni ja ksantofülli). Nende värvus varieerub kollasest (kontlikroonlehtedes) ja oranžist (oranžikestes) kuni oranžikaspunase (porgandijuurtes) ja erkpunase (kibuvitsamarjade) värvini.

Riis. 33. Nasturtiumi tuppleherakk Tropaeolum majus tuuma ja kromoplastidega.

1 - naharakkudes Philodendron grandifolium; 2, 3- seemnerakkudes Melandrium macrocarpum; neli- juurerakkudes Phajus grandifolius(näidatud on ainult tuum, mille läheduses on leukoplastid).
Kromoplastide kuju on väga mitmekesine. Mõnikord on need ellipsoidsed või, kui need on tihedalt asetsevad, hulknurksed tabelikujulised, labakujulised jne. Tavaliselt on kromoplastil nõelakujuline ja nurgeline kuju, selle stroom on venitatud pigmendiga, mis moodustab kromoplasti põhiosa (joonis 33). Paljude taimede kromoplastides on tärklist.
Mõnikord eraldatakse kromoplastides valkaineid või tekivad õlitilgad: apelsiniviljade ja teiste tsitrusviljade koore rakkudes on kromoplastide pigment osaliselt lahustunud eeterlikes õlides.
Viljade värvus ei sõltu alati ainult kromoplastidest. Plastiidide olemasolu määrab värvid rohelise, kollase, telliskivipunase (kontlillõie kroonlehed, Compositae, porgandijuured, paljud valmivad viljad). Sinise, vaarika, tumepunase (vaarika, viburnumi viljad) värvused sõltuvad antotsüaniine sisaldava rakumahla värvusest. Sageli saadakse rakumahla ja plastiidide värvuse tõttu segavärv.
Väga oluline on karoteeni eraldamine benseeniga punase pipra ja pihlaka viljade alkoholiekstraktidest. Kui pipraviljade alkoholiekstraktile lisatakse benseeni, lahustub karoteen benseenis peal ja ksantofüll on alkoholis allpool. Rakumahl (jääb ka piiritusse) on värvitu. Kui korrata seda katset pihlakaviljadega, on näha, et neis pole nii palju karoteeni, kuna benseenifraktsioon on palju kahvatum kui punase pipra oma ja alkohol jääb antotsüaniini olemasolust roosaks.
Kroonlehtede valge värvus on tingitud valguse ühtlasest hajumisest, kui see peegeldub kõrgelt arenenud rakkudevahelistes ruumides ja elundi pinnal asuvatest õhumullidest.
Mõnede õite ja viljade (daalia, kärbseseen, moon, sidrun) kollane värvus sõltub ka rakumahla pigmendist, mis on seotud antotsüaniiniga – antoklooriga.
Taimeosade värvus võib sõltuda ka surnud rakkude värvist ja nende muutunud sisust (seemnekest, puutüvede pind).
Leukoplastid. Plastiidid, mis ei sisalda stroomas pigmente ja mida nimetatakse leukoplastideks, esinevad enamiku taimede paljudes rakkudes (joonis 34). Kuna leukoplastid on värvitud ja pealegi murravad valgust samamoodi nagu protoplasmid, ei ole nende olemasolu rakus alati lihtne tuvastada.
Leukoplastid on tavaliselt peaaegu sfäärilise kujuga. Juhtudel, kui nende stroomas leidub tärklist või valku, omandavad nad teistsuguse kuju. Hariduskuded, maa-alused elundid ja seemned on suhteliselt rikkad leukoplastide poolest.
Leukoplastides võib moodustuda tärklis, mis ladestub terade kujul nende stroomas.
Paljudel juhtudel koguneb tärklist leukoplasti nii suures koguses, et plastiidi (strooma) eluskeha surutakse perifeeriasse. Seda võib mõne raskusega näha väga õhukese kilena tärklisetera pinnal; nendel juhtudel on leukoplast tärklise akumulaator täies mahus (tärklise varu kohta vt lk 78).
plastiidi liikumine. Lisaks plastiidide liikumistele, mis on seotud nende jaotumisega rakkude jagunemise tulemusena tekkinud tütarrakkude vahel, toimuvad muud liikumised: 1) sõltuvalt vanusega seotud muutustest rakkudes ja plastiidides ning 2) pöörduvad ja korduvalt korduvad liikumised, mis on seotud muutustega. keskkonnateguritega kokkupuute suunas ja intensiivsuses (plastiidide "ligikaudsed" liikumised).
Protoplasma seinakihis asuvad klorofülliterad võivad rakus liikuda. Protoplasma ei kanna neid mitte ainult passiivselt, kui see liigub, vaid sõltuvalt valguse tugevusest ja suunast võivad nad ka ise oma asukohta rakus muuta. I.P. Borodin näitas, et see on kõige paremini nähtav pardlille õhukestel lehtedel ( Lemna trisulca).
Joonisel 35 on näidatud klorofülli terade paiknemine rakkudes, mis olid erinevates valgustingimustes. Hajuvalguses paiknevad klorofülli terad piki seinu, mis on kiirte põhisuunaga täisnurga all; suurem osa langevast valgusest tabab otse iga tera. Ereda päikesevalguse käes liiguvad klorofülli terad külgseintele, mis asuvad
mööda langevaid kiiri. Rakku tungiv valgus ei taba otse tera, vaid hajub külgedele ja valgustab terad ühtlasemalt. Pimedas asuvad terad mööda seinu, mis puutuvad kokku naaberrakkude seintega, nagu öeldakse soojadel seintel, kuid viimast paigutust pole alati võimalik jälgida.
Huvitaval kombel toimub üksikute klorofülliterade valgustus väga ühtlaselt. Klorofülli terade iseseisev liikumiskiirus rakus on 0,12 μ sekundis, st kiirus on palju väiksem kui amööbil ja plasmoodiumil.
Klorofülli terade asukohta ei mõjuta mitte ainult valguse tugevus ja suund. Mõnel juhul täheldati klorofülli terade liikumist nendesse rakuseina osadesse, kuhu sobivad rakkudevahelised ruumid; süsihappegaas, mis imendub assimilatsiooniprotsessis, siseneb rakkudevahelistesse ruumidesse.

Riis. 35. Klorofülli terade liikumine pardipuu trikoo rakkudes ( Lemna trisulca):
AGA- osa taimest plaanis, ülemine vasak osa oli pimeduses, alumine parem osa hajusvalguses (Borodini järgi); B- vanandatud taimede osalised põikilõiked: 1 - hajutatud valguses; 2 - otsese päikesevalguse käes; 3 - Pimedas.
Plastiidide päritolu. Erinevate kategooriate plastiidide vahel pole teravaid piire. Ühte tüüpi plastiidid võivad muutuda teist tüüpi plastiidideks. Tõepoolest, seemikute ja täiskasvanute puhul muutuvad paljud embrüonaalsete rakkude leukoplastide jagunemisel tekkivad plastiidid kloro- ja kromoplastideks. Kloroplast, kaotades klorofülli ja olles rikastatud karotenoididega (nagu valmivate kibuvitsade puhul), muutub kromoplastiks; leukoplast, kui karotenoidid kogunevad selle stroomas, muutub kromoplastiks (nagu porgandijuurtes). Piisab mullas kasvava porgandi punase "juurvilja" ülemise osa paljastamisest maapinnast, nii et mõne aja pärast muutub see hüpokotüüli ja juure osa roheliseks: rakkude kromoplastid muutuvad kloroplastideks.
Küsimus plastiidide esialgsest ilmumisest taimerakkudesse ei ole praegu veel täiesti selge. Oli teooria plastiidide päritolu kohta kondriosoomidest; enamik teadlasi on sellest loobunud. Viimasel ajal on avaldatud arvamust, et plastiidid tekivad spetsiaalsetest moodustistest - proplastiididest.
Praegu on teada, et klorofüll esineb taimedes mitmel kujul, kuid teada on ainult kahe selle vormi struktuur - a ja b, ja ka siin tuleb veel selgitada kaksiksidemete asukoht ning magneesiumi ja lämmastiku vahelise sideme olemus.
A. S. Famintsyn tegi esimesena kindlaks, et fotosünteesi protsess võib toimuda ka kunstliku valgustuse korral.
Need on rühmitatud granadeks, mis on lamestatud ja tihedalt üksteise külge surutud kettakujuliste tülakoidide virnad. Grana ühendatakse lamellide abil. Kloroplasti membraani ja tülakoidide vahelist ruumi nimetatakse stroomiks. Stroma sisaldab kloroplasti RNA molekule, plastiidi DNA-d, ribosoome, tärkliseterasid, aga ka Calvini tsükli ensüüme.
Päritolu
Kloroplastide sümbiogeneesi päritolu on nüüdseks üldtunnustatud. Eeldatakse, et kloroplastid pärinevad tsüanobakteritest, kuna need on kahe membraaniga organoidid, neil on oma suletud ringikujuline DNA ja RNA, täielik valgusünteesiaparaat (pealegi prokarüootset tüüpi ribosoomid - 70S), paljunevad binaarse lõhustumise teel ja tülakoidmembraanid on sarnased prokarüootsete membraanidega (happeliste lipiidide olemasolu) ja sarnanevad sinivetikate vastavate organellidega. Glaukofüütvetikates on tüüpiliste kloroplastide asemel rakkudes tsüanella - tsüanobakterid, mis on endosümbioosi tagajärjel kaotanud iseseisva eksisteerimise võime, kuid säilitanud osaliselt tsüanobakteri rakuseina.
Selle sündmuse vanuseks hinnatakse 1–1,5 miljardit aastat.
Mõned organismirühmad said kloroplaste endosümbioosi tulemusena mitte prokarüootsete rakkudega, vaid teiste eukarüootidega, millel on juba kloroplastid. See seletab rohkem kui kahe membraani olemasolu mõne organismi kloroplasti membraanis. Nendest membraanidest sisemist tõlgendatakse rakuseina kaotanud tsüanobakteri kestana, välimist aga peremeesorganismi sümbiontofoori vakuooli seinana. Vahemembraanid - kuuluvad sümbioosi astunud redutseeritud eukarüootse organismi hulka. Mõnes rühmas on teise ja kolmanda membraani vahelises periplastiidi ruumis nukleomorf, tugevalt redutseeritud eukarüootne tuum.
Kloroplasti mudel
Struktuur
Erinevates organismirühmades erinevad kloroplastid oluliselt nii suuruse, struktuuri kui ka arvu poolest rakus. Kloroplastide struktuuri tunnused omavad suurt taksonoomilist tähtsust.
Kloroplastide kest
Erinevates organismirühmades on kloroplastide kest ehituselt erinev.
Glaukotsüstofüütidel, punastel, rohelistel vetikatel ja kõrgematel taimedel koosneb kest kahest membraanist. Teistes eukarüootsetes vetikates on kloroplast lisaks ümbritsetud ühe või kahe membraaniga. Neljamembraanilise kloroplastiga vetikatel ulatub välismembraan tavaliselt tuuma välismembraani.
Periplastiidne ruum
Lamell ja tülakoidid
Lamellid ühendavad tülakoidide õõnsusi.
Pürenoidid
Pürenoidid on polüsahhariidide sünteesi keskused kloroplastides. Pürenoidide struktuur on mitmekesine ja need ei ole alati morfoloogiliselt väljendunud. Need võivad olla intraplastid ja varred, ulatudes tsütoplasmasse. Rohevetikates ja taimedes paiknevad pürenoidid kloroplasti sees, mis on seotud tärklise intraplastiidse ladustamisega.
Stigma
Stigmasid või silmi leidub liikuvate vetikarakkude kloroplastides. Asub flagellumi aluse lähedal. Stigmad sisaldavad karotenoide ja on võimelised toimima fotoretseptoritena.
Vaata ka
Märkmed
Kommentaarid
Märkmed
Kirjandus
- Belyakova G. A. Vetikad ja seened // Botaanika: 4 köites / Belyakova G. A., Dyakov Yu. T., Tarasov K. L. - M.: Kirjastuskeskus "Akadeemia", 2006. - T. 1. - 320 lk - 3000 eksemplari. - ISBN 5-7695-2731-5
- Karpov S.A. Protisti raku struktuur. - Peterburi. : TESSA, 2001. - 384 lk. - 1000 eksemplari. - ISBN 5-94086-010-9
- Lee, R.E. Füsioloogia, 4. väljaanne. - Cambridge: Cambridge University Press, 2008. - 547 lk. - ISBN 9780521682770
| eukarüootsete rakkude organellid | ||
|---|---|---|
 |
||
Wikimedia sihtasutus. 2010 .
Vaadake, mis on "kloroplastid" teistes sõnaraamatutes:
- (kreeka keelest chloros green ja plastos fashioned), taimede rakusisesed organellid (plastiidid), milles toimub fotosüntees; klorofülli tõttu on need värvunud roheliseks. Leitud rakkudes taimede maapealsete elundite kuded, ... ... Bioloogia entsüklopeediline sõnastik
- (kreeka keelest chloros green ja plastos fashioned moodustuvad), taimeraku rakusisesed organellid, milles toimub fotosüntees; on värvitud roheliseks (need sisaldavad klorofülli). Oma geneetiline aparaat ja ...... Suur entsüklopeediline sõnaraamat
Taimerakkudesse suletud kehad, mis on värvitud roheliseks ja sisaldavad klorofülli. Kõrgemate taimede klorofüllid on väga kindla kujuga ja neid nimetatakse klorofülli teradeks; vetikates on nende kuju mitmekesine ja neid nimetatakse kromatofoorideks või ... Brockhausi ja Efroni entsüklopeedia
Kloroplastid- (kreeka keelest chloros green ja plastos fashioned, moodustunud), taimeraku rakusisesed struktuurid, milles toimub fotosüntees. Need sisaldavad pigmenti klorofülli, mis annab neile rohelise värvi. Kõrgemate taimede rakus 10 kuni ... Illustreeritud entsüklopeediline sõnaraamat
- (gr. klororoheline + laste moodustamine) taimeraku rohelised plastiidid, mis sisaldavad klorofülli, karotiini, ksantofülli ja osalevad fotosünteesi protsessis vrd. kromoplastid). Uus võõrsõnade sõnastik. EdwART, 2009. kloroplastid [gr.… … Vene keele võõrsõnade sõnastik
- (kreekakeelsetest sõnadest chlorós green and plastós fashioned, moodustunud) Plastidi taimeraku rakusisesed organellid, milles toimub fotosüntees. Need on värvitud roheliseks, kuna neis on fotosünteesi põhipigment ... Suur Nõukogude entsüklopeedia
Ov; pl. (ühik. kloroplast, a; m.). [kreeka keelest. chlōros kahvaturohelised ja vormitud plastos] Nerd. Taimerakkude protoplasmas olevad kehad, mis sisaldavad klorofülli ja osalevad fotosünteesi protsessis. Klorofülli kontsentratsioon kloroplastides. * * *…… entsüklopeediline sõnaraamat
Taimerakkudesse suletud kehad, mis on värvitud roheliseks ja sisaldavad klorofülli. Kõrgematel taimedel on X. väga kindla kujuga ja neid nimetatakse klorofülli teradeks (vt.); vetikates on nende kuju mitmekesine ja neid nimetatakse ... ... Entsüklopeediline sõnaraamat F.A. Brockhaus ja I.A. Efron
Mn. Taimeraku rohelised plastiidid, mis sisaldavad klorofülli, karoteeni ja osalevad fotosünteesi protsessis. Efraimi selgitav sõnaraamat. T. F. Efremova. 2000... Kaasaegne vene keele seletav sõnaraamat Efremova
- (kreeka keelest chloros green ja plastos moodustunud, moodustunud), kasvavad rakusisesed organellid. rakud, milles toimub fotosüntees; on värvitud roheliseks (need sisaldavad klorofülli). Omad geneetiline aparatuur ja valkude süntees ...... Loodusteadus. entsüklopeediline sõnaraamat
Plastiidid on protoplasti organellid, mis on iseloomulikud ainult taimerakkudele. Need puuduvad ainult bakterites, sinivetikates ja võib-olla ka seentes.
Kõrgemates taimedes leidub plastiide kõigi elundite täiskasvanud vegetatiivsetes rakkudes – varres, lehtedes, juurtes ja õites. Plastiidid on suhteliselt suured organellid, mis on palju suuremad kui mitokondrid ja mõnikord isegi suuremad kui tuum, tihedamad kui neid ümbritsev tsütoplasma, mis on valgusmikroskoobis selgelt nähtavad. Neil on iseloomulik struktuur ja nad täidavad mitmesuguseid funktsioone, mis on peamiselt seotud orgaaniliste ainete sünteesiga.
Täiskasvanud taimerakus eristatakse sõltuvalt värvist, kujust ja funktsioonist kolme peamist plastiidide tüüpi: kloroplastid (rohelised plastiidid), kromoplastid (kollased ja oranžid plastiidid) ja leukoplastid (värvitud plastiidid). Viimased on mõõtmetelt väiksemad kui kahe eelneva tüübi plastiidid.
Kloroplastid
Kloroplasti struktuurne alus on valgud (umbes 50% kuivkaalust), need sisaldavad ka 5-10% klorofülli ja 1-2% karotenoide. Nagu mitokondrites, leiti ka kloroplastides väike kogus RNA-d (0,5–3,5%) ja veelgi vähem DNA-d. Kloroplastide erakordne tähtsus seisneb selles, et nad viivad läbi fotosünteesi protsessi. Fotosünteesi käigus tekkinud tärklist nimetatakse primaarseks ehk assimilatsiooniks, see ladestub kloroplastidesse väikeste tärkliseteradena. Klorofülli olemasolu on vajalik normaalseks fotosünteesi kulgemiseks. Klorofüll on fotosünteesi elluviimisel peamine toimeaine. See neelab valguse energiat ja suunab selle fotosünteetiliste reaktsioonide läbiviimiseks. Klorofülli saab plastiididest ekstraheerida alkoholi, atsetooni või muude orgaaniliste lahustitega. Kollaste pigmentide rolli fotosünteesis ei mõisteta ikka veel hästi. Eeldatakse, et nad neelavad ka päikeseenergiat ja kannavad selle üle klorofüllile või viivad koos sellega läbi spetsiifilisi fotosünteesi jaoks olulisi reaktsioone.
Vastavalt oma funktsioonidele paiknevad kloroplastid peamiselt valguse poole jäävates fotosünteetilistes elundites ja kudedes – lehtedes ja noortes vartes, valmimata viljades. Mõnikord leidub kloroplaste isegi juurtes, näiteks maisi juhuslikes juurtes. Kuid enamik neist on koondunud lehe mesofülli (pulbi) rakkudesse.
Erinevalt teistest organellidest iseloomustab kõrgemate taimede kloroplaste kuju ja suuruse ühtlus ning püsivus. Need on enamasti kettakujulised või läätsekujulised ning lamades on ümarad või hulknurksed. Sel juhul nimetatakse neid sageli ka klorofülli terad. Kloroplastide suurus on üsna konstantne ja isegi erinevatel kõrgemate taimede liikidel varieerub veidi, keskmiselt 3-7 mikronit (paksus 1-3 mikronit). Suuremad kloroplastid on kõrgemates taimedes haruldased. Näiteks Selaginellas (lükopoodid) leidub lehtede naharakkudes üks või kaks suurt lamellkloroplasti. Kloroplastide suurus ja kuju muutuvad sõltuvalt välistingimustest. Varju armastavates taimedes on kloroplastid üldiselt suuremad kui valgust armastavatel taimedel ja reeglina klorofüllirikkamad. Tavaliselt kannab rakk suurt hulka kloroplaste ja nende arv on väga erinev; keskmiselt on selles 20 kuni 50 kloroplasti. Lehed on eriti rikkad kloroplastide, aga ka noorte valmimata viljade poolest. Kloroplastide koguarv taimes võib olla tohutu; näiteks täiskasvanud puus on kümneid ja sadu miljardeid kloroplaste. Kloroplastide arv rakus on seotud nende suurusega. Nii sisaldavad maisi leherakud tavaliselt mitut kloroplasti, kuid eriti suurte kloroplastidega sortidel väheneb nende arv raku kohta kahele.
Paljudes madalamates taimedes (vetikates) on kloroplastide kuju, arv ja suurus väga mitmekesised. Need võivad olla lamelljad (Mougeotia), tähekujulised (Zygnema) või spiraalsete paelte (Spirogyra) ja ribiliste silindrite kujul (Closterium). Sellised kloroplastid on tavaliselt väga suured, neid esineb rakus vähe (ühest mitmeni) ja neid nimetatakse kromatofoorid. Kuid isegi vetikates võib leida tavalise läätsekujulise kujuga kloroplaste ja sel juhul on nende arv rakus tavaliselt suur.
Kõrgemate taimede rakkudes paiknevad kloroplastid tsütoplasmas nii, et nende üks lame külg jääb rakumembraani poole ja eriti palju on neid õhuga täidetud rakkudevaheliste ruumide läheduses. Siin külgnevad nad tihedalt üksteisega ja nende piirjooned muutuvad nurgeliseks. Kloroplastide asend rakus võib aga muutuda sõltuvalt välistingimustest ja eelkõige valgustusest. Need asuvad puuris nii, et jäädvustavad valgust parimal viisil, ilma et nad samal ajal otsese päikesevalguse käes oleksid. Mõnede taimede lehtedes hajutatud valguses paiknevad kloroplastid peamiselt nendel rakumembraani seintel, mis on suunatud elundi pinna poole, eredas valguses aga koonduvad külgseintele või pöörduvad kitsa küljega kiirte poole, st serv. Mõnikord täheldatakse samasugust kloroplastide liikumist ka muude stiimulite – temperatuuri, keemilise, mehaanilise jne – mõjul. Kas plastiidide liikumine on aktiivne või passiivne (tsütoplasmaatiline vool), pole veel täielikult välja selgitatud, kuid praegu on argumente rohkem aktiivse liikumise kasuks.
Fotosünteesi protsesside keerukuse põhjal, mis koosnevad paljudest reaktsioonidest, millest igaüht katalüüsib konkreetne ensüüm, võib eeldada, et kloroplastidel on korrastatud ja keeruline struktuur. Tõepoolest, isegi tavalises valgusmikroskoobis on sageli näha, et kloroplastid ei ole täiesti homogeensed, vaid neis on täheldatud tumedamaid väikeseid terakesi, mis on orienteeritud paralleelselt kloroplasti pinnaga, mis said nime. terad. Elektronmikroskoobiga tehtud uuringud kinnitasid granaadi olemasolu ja näitasid, et kogu kloroplastil tervikuna ja graanal on keeruline struktuur.
Nagu mitokondrid, on kloroplastid membraanistruktuurid, mis asuvad vabalt tsütoplasmas. Need on tsütoplasmast piiritletud kahemembraanilise membraaniga, mille membraanide vahel on selgelt nähtav valgusvahe. Arvatakse, et membraanid on siledad ega sisalda kinnitunud osakesi. Kuni viimase ajani arvati, et kloroplasti membraan on pidev, sellel ei olnud auke ja selle membraanid ei ole ühendatud endoplasmaatilise retikulumi membraanidega. Kuid nüüd koguneb tõendeid, mis näitavad, et see pole alati nii. Mõnikord võivad kesta mõnes kohas tekkida submikroskoopilised augud. Võimalik, et teatud kloroplasti aktiivsuse perioodidel on need "poorid" tihedas kontaktis endoplasmaatilise retikulumiga, kuid see kontakt on lühiajaline. Kloroplasti membraan, millel on selektiivne läbilaskvus, mängib tsütoplasma ja kloroplasti vahelises metabolismis reguleerivat rolli.
Kloroplasti keha läbistab kahe membraaniga plaatide süsteem nn lamellid. Lamellide vaheline ruum on täidetud vesise valguvedelikuga - strooma või kloroplasti maatriks. Stroma võib sisaldada tärkliseterasid, õlitilku ja ribosoomitaolisi osakesi. Üsna hiljuti on mõnede taimede kloroplastide stroomas eriti peeneid valmistamismeetodeid kasutades leitud 80–100 Å läbimõõduga ja üle 1000 Å pikkuste paralleelsete fibrillide klastreid. Neid kloroplasti mikrofibrillide kimpe nimetatakse stromakeskused. Nende funktsioon pole täiesti selge.
Mõnes kloroplasti piirkonnas haakuvad lamellid üksteisega üsna tihedalt, olles selle pinnaga paralleelsed, mille tulemusena tekivad nendes piirkondades lamellide kobarad, mida nimetatakse granaks. Grana sees liituvad servades paaritud lamellmembraanid, moodustades kinnised lapikkotid nn. plaadid, või tülakoidid. Selliste ketaste pakid moodustavad granaadi. Eraldi tahud ühendatakse liideseruumi läbivate lamellide abil ühtseks süsteemiks. Klorofüll ei ole hajusalt hajutatud kogu kloroplastis, vaid kontsentreerub lamellidesse, nagu eeldatakse, monomolekulaarse kihina. Ribosoome ei leidu mitte ainult maatriksis, vaid neid võib leida ka graniidi pinnalt.

Terades olevate ketaste arv ulatub kahest kuni mitmekümneni ja läbimõõt on olenevalt taime tüübist 0,3 kuni 2 mikronit. Seetõttu pole paljudes taimedes grana valgusmikroskoobi all nähtav. Granaatide arv ja paigutus kloroplastis oleneb taime tüübist, kloroplastide vanusest ja aktiivsusest. Aspidistra kloroplastides on graniidid nii palju, et need on üksteisega kontaktis ja kõrvitsa niinimetatud satelliitrakkudes hõivab kloroplasti põhimahu strooma. Tomati ja krüsanteemi lehtede kloroplastides on granad hajutatud juhuslikult, tubaka kloroplastides on need aga kloroplasti pinna suhtes õigesti orienteeritud ja asuvad üksteisest võrdsel kaugusel. Valguslembestel taimedel on terad väiksemad kui varjutaluvatel.
Kõrgemate taimede kloroplastide struktuur on suurepäraselt kohandatud nende põhifunktsiooni - fotosünteesi - täitmiseks. Juba klorofülli kandva aparaadi jagunemine väikesteks plastiidideks tähendab aktiivse pinna tohutut suurenemist. Tänu membraanide ja terade moodustumisele suureneb see pind veelgi. Suur aktiivne pind ja peen ruumiline orientatsioon tagavad lihtsa juurdepääsu valguskvanti energiale ja võimaluse seda energiat üle kanda fotosünteesis osalevatele keemilistele süsteemidele. Suletud kambrite põhimõte - tülakoidid võimaldavad ruumilise eraldatuse tõttu samaaegselt ja iseseisvalt läbi viia sama reaktsioonide kompleksi, mis moodustavad fotosünteesi. Valkude süntees toimub kloroplastide ribosoomides.
Mõnede vetikate (spirogyra) ja harvemini kõrgemate taimede (näiteks maisi nn veresoonte kimpude voodri rakud) rakkudes leitakse näota kloroplastid, milles lamellid tungivad läbi stroomas, moodustamata selgelt eristuvaid omadusi. graanulid.
Kloroplastide päritolu ja arengut on uuritud väga vähe ning ühest seisukohta selles küsimuses veel ei ole. On teada, et noortes embrüonaalsetes rakkudes ei esine diferentseerunud kloroplaste. Selle asemel on nö proplastiidid. Need on väga väikesed (mikroniosakesed) kehad, mis on valgusmikroskoobi lahutusvõime piiril. Esialgu on neil amööboidne kuju (kandesagarad), need on tsütoplasmast piiritletud topeltmembraaniga ja ei sisalda ei sisemembraane ega klorofülli. Lamelle moodustavad sisemembraanid arenevad hiljem. Proplastiidide edasise arengu kohta on mitmeid hüpoteese. Neist ühe järgi tekivad proplastiidide läbipaistvas stroomas esmalt pisikeste mullide kobarad, mis on paigutatud õigesse järjekorda nagu kristallvõre. Seda mullide kogunemist, millest igaüks on kaetud oma membraaniga, nimetatakse esmane tahk. Mööda primaarse grana perifeeriat tekivad lamellid, mis levivad sellest igas suunas. Seejärel moodustuvad neile kõik kloroplasti lamellstruktuurid, sealhulgas grana. Valguses näitavad need pigmentide ja eelkõige klorofülli ladestumist.
Teise hüpoteesi kohaselt moodustuvad lamellid algselt proplastiidi ümbrise sisemembraani voltidena, mitte vesiikulitest. Sel juhul ilmneb esmalt mitokondritega sarnane struktuur.
Seega lähtuvad need hüpoteesid plastiidi järjepidevuse printsiibist ja eitavad oma päritolu teistest protoplasti organellidest ja eelkõige tsütoplasmast. Teised teadlased usuvad aga, et mitokondrid ja plastiidid on päritolult tihedalt seotud. Näiteks oli võimalik näidata mitokondrite tekkimist küpsetest kloroplastidest "pungamise teel". Seejärel võisid need mitokondrid rekombineeruda kloroplastidega. Kuid kõik need hüpoteesid pole veel piisavalt põhjendatud ja plastiidide päritolu küsimus ootab endiselt otsustamist.
Lisaks sellele, et kloroplastid pärinevad proplastiididest, võivad nad paljuneda ka lihtsa jagunemise teel. Sel juhul moodustub täiskasvanud kloroplastist kaks tütarplastiidi, sageli ebavõrdse suurusega. Sellise lõhustumise elektronmikroskoopilist pilti pole veel uuritud.
Kloroplasti struktuur ei jää konstantseks, see muutub loomulikult rakkude kasvuprotsessis. Kloroplastide struktuuri muutus lehtede vanusega on märgatav isegi valgusmikroskoobis. Seega vastavad noored lehed tavaliselt peeneteralisele struktuurile, keskealised lehed jämedateralisele struktuurile. Vananevates lehtedes on struktuur häiritud ja kloroplastid lagunevad.
Kloroplastid on üsna õrnad haprad organellid. Kui rakud asetada destilleeritud vette või hüpotoonilisse soolalahusesse, paisuvad need kiiresti, nende pinnale tekivad mullilaadsed tursed ja seejärel levivad. Elektronmikroskoopilised uuringud on näidanud, et turse esineb stroomas, mitte lamellides. Kui rakk on kahjustatud, muutuvad tavapärase mikroskoobiga vaadeldavad kloroplastid esmalt jämedalt teraliseks, paisuvad, omandavad vahuse välimuse ja lõpuks kaob granulaarsus. Patoloogilised muutused tekivad lehtede kloroplastides ja mineraalsete toitainete puudumisega mullas. Siiski võivad mõnede rakkude kloroplastid näidata ka suurt vastupidavust. Niisiis, puudel on koore roheline värvus tingitud kloroplastidega rakukihi olemasolust. Need kloroplastid taluvad suurepäraselt madalaid temperatuure ja lähevad aktiivsesse olekusse, mille tuvastab koore tugev haljendamine, näiteks haabjas, väga varakevadel, kui öösel on veel tugevad külmad. Talvist madalat temperatuuri taluvad ka meie igihaljaste okaspuude lehtede (okaste) kloroplastid. Samal ajal, nagu näitavad elektronmikroskoopilised uuringud, säilitavad nad oma keeruka sisemise korralduse.
Leukoplastid
Need on väikesed värvitud plastiidid. Valgusmikroskoobis on neid sageli raske tuvastada, kuna need on värvitud ja neil on sama murdumisnäitaja kui tsütoplasmal. Neid saab tuvastada ainult siis, kui nende sisse kogunevad suured kandmised. Need on väga õrnad organellid ja elusmaterjali lõikude valmistamisel hävivad need isegi kergemini kui kloroplastid. Neid leidub päikesevalguse eest varjatud täiskasvanud rakkudes: juurtes, risoomides, mugulates (kartulites), seemnetes, varresüdamikes ja ka tugeva valgusega kokkupuutuvates rakkudes (naharakud). Leukoplastid kogunevad sageli tuuma ümber, mõnikord ümbritsevad seda igast küljest. Leukoplastide kuju on väga varieeruv, enamasti on need sfäärilised, munajad või spindlikujulised moodustised.

Leukoplastid on organellid, mis on seotud varutoitainete – tärklise, valkude ja rasvade – moodustumisega. Leukoplastide aktiivsus on spetsialiseerunud: mõned neist koguvad peamiselt tärklist (amüloplastid), teised - valgud (proteoplastid, mida nimetatakse ka aleuronoplastideks) ja teised - õlid (oleoplastid). Lehtede ja varte naharakkude leukoplaste ei saa ühelegi neist tüüpidest omistada, kuna nende funktsiooni pole veel selgitatud.
Amüloplastid koguma tärklist nn tärkliseterade kujul. See on domineeriv leukoplasti tüüp. Amüloplastide struktuuri ja tärklise moodustumise mehhanismi on valgusmikroskoobis raske uurida ning elektronmikroskoobis on neid veel vähe uuritud. Eeldatakse, et need on moodustunud proplastiididest, kuid erinevalt kloroplastidest ei jõua nende struktuuri areng kaugele, vaid viibib üsna varajases staadiumis - ebaküpse, nõrgalt lamellaarse plastiidi staadiumis. Väljaspool on amüloplastid piiritletud kahe membraaniga membraaniga. Plastiidi sees on täidetud peeneteralise stroomaga. Tärkliseterade tekkele amüloplastides eelneb pisikeste vesiikulite teke, mis ühinevad, lamenduvad, piirates membraaniga stroomaala plastiidi keskel. See piirkond, nimega hariduskeskus, muutub heledamaks, meenutades vakuooli. Hariduskeskuses algab tärklise ladestumine. Kui tulevane tärklisetera suurus hakkab suurenema, kaovad hariduskeskust piiritlevad membraanid ja edasine terade kasv läheb neist kaugemale. Tärklise ladestamisel võivad amüloplasti ümbris ja strooma tugevasti venitada, mille tulemusena suureneb tärklise tera kasvu tõttu tugevasti amüloplasti suurus. Tärklisetera täidab seejärel peaaegu kogu amüloplasti õõnsuse, surudes selle elava sisu väga õhukese kile kujul tera pinnale perifeeriasse. Paljudel juhtudel võib tärklisetera saavutada sellised mõõtmed, et amüloplast puruneb ja jääb tärklisetera vaid ühele küljele. Sel juhul saab uusi tärklise portsjoneid sünteesida ainult nendes piirkondades, kus tärklisetera jääb kontakti amüloplasti membraanide ja stroomaga.
Proplastiididest arenedes võivad amüloplastid teatud tingimustel muutuda teist tüüpi plastiidideks. Kui näiteks odrajuur asetada valguse kätte, on näha, et osa leukoplastidest suureneb mahult ja muutuvad sarnaselt lehtedes tekkivateks kloroplastideks. Kui selline juur jääb jälle valgusest ilma, siis need kloroplastid vähenevad ja kaotavad oma klorofülli, kuid ei muutu tagasi leukoplastideks, vaid toodavad karoteeni kerakesi (pallikesi), muutudes nii kromoplastideks. Oleoplastid st leukoplastid, mis moodustavad peamiselt õlisid, on palju vähem levinud kui amüloplastid (näiteks mõne üheidulehe lehtede rakkudes). Need on tavaliselt vananevate kloroplastide saadused, mis kaotavad klorofülli. Sel juhul ilmuvad plastiidi stroomas väikseimad õlikuulikesed. Seejärel plastiidi membraan laguneb ja naaberplastiidide sisu ühineb, moodustades suuremad rasvatilgad. Mõnikord koguneb sellistesse plastiididesse korraga ka tärklis.
Säilitusvalgu - valgu - sünteesi saab läbi viia kolmandat tüüpi leukoplastides - proteoplastid. Valk kristallide ja teradena moodustub paljude taimede seemnetes, eriti nendes, mis sisaldavad ka palju õli (näiteks kastoorseemned). Proteoplastid, nagu amüloplastid, tekivad proplastiididest. Nende areng hilineb ka ebaküpse lamellplastiidi staadiumis. Proteoplasti stroomas koguneb säilitusvalk algul fibrillide kujul, mis seejärel ühinevad suuremateks kimpudeks. Lisaks hävivad plastiidi kest ja strooma ning valgufibrillide kimbud muutuvad omamoodi väikesteks viskoosseteks vakuoolideks. Seejärel ühinevad naaberplastiidide vakuoolid, osa valgust moodustub kristalloidide kujul.
Seega on nii tärklis, säilitusvalgud kui ka õlitilgad inertsed kandmised, plastiidide jääkproduktid. Pealegi võib igaüks neist akumuleeruda mitte ainult leukoplastides, vaid ka kloroplastides ja kromoplastides. Aga kui tärklis tekib plastiidides, siis säilitusvalgud ja -rasvad võivad väga sageli olla ka ekstraplastiidse päritoluga, tõenäoliselt tekivad otse tsütoplasmas ja plastiididest sõltumatult. Sel juhul toimuvad struktuuriprotsessid on endiselt halvasti mõistetavad.
Kromoplastid
Kromoplastid on kollased või oranžid ja isegi punased plastiidid. Neid leidub paljude kroonlehtede (võilill, ranunculus, saialill), küpsete viljade (tomatid, metsroos, pihlakas, kõrvits, arbuus, apelsin), juurviljade (porgand, söödapeet) rakkudes. Nende elundite ere värvus on tingitud kollastest ja oranžidest pigmentidest - karotenoididest, mis on koondunud kromoplastidesse. Need pigmendid on iseloomulikud ka kloroplastidele, kuid seal varjab neid klorofüll. Need on vees lahustumatud, kuid lahustuvad rasvades.
Erinevalt kloroplastidest on kromoplastide kuju väga muutlik ja selle määrab nende päritolu ja pigmentide olek neis, samuti neid moodustava taime süstemaatiline asend. Sõltuvalt karotenoidide seisundist eristatakse kolme tüüpi kromoplaste:
- kromoplastid, milles karotenoidid ladestuvad väikeste, kuid omavahel mitteseotud kristallidena, mis on valgusmikroskoobi all nähtavad (porgandikromoplastid);
- kromoplastid, milles karotenoidid on lahustunud submikroskoopilistes lipoidkerakestes (buttercup kroonlehed ja aaloe);
- kromoplastid, mille karotenoidid on kogutud kimpudesse, mis koosnevad submikroskoopilistest filamentidest ja on seotud valgufibrillidega (punane pipar, tomat, mandariin).
Erinevalt kloroplastidest ja leukoplastidest tekivad kromoplastid harva otse proplastiididest, kuid on tavaliselt kloroplastide degeneratsiooni tagajärg. Erandiks on porgandikromoplastid, mis tekivad mitte kloroplastidest, vaid leukoplastidest või otse proplastiididest. Need juurvilja osad, mis ei ole mulda kastetud ja arenevad valguse käes, muutuvad tavaliselt roheliseks. See ei toimu kromoplastide kloroplastideks muutumise tulemusena, vaid kloroplastide moodustumise tõttu proplastiididest või leukoplastidest. Kromoplastid ei saa üldse muutuda teist tüüpi plastiidideks. Kõige sagedamini tekivad kromoplastid kloroplastide hävitamise käigus, kui viimased satuvad pöördumatusse arengufaasi. See on 2. ja 3. tüüpi kromoplastide päritolu. Samal ajal suureneb rasvade ja karotenoidide sisaldus kloroplastides, mis kogunevad plastiidi stroomas submikroskoopiliste gloobulite kujul, kaovad lamellstruktuurid, hävib klorofüll. Pigmendi gloobulid kasvavad ja strooma maht väheneb, mille tulemusena suudavad gloobulid täita suurema osa plastiidist. "Ema" kloroplasti ümar kuju on säilinud. Sarnane kloroplastide lagunemisprotsess toimub tõenäoliselt lehtede sügisene kollaseks muutumise ja viljade valmimise ajal. Koltuvates lehtedes sisalduv klorofüll hävib ja ei varja enam karotenoide, mis eenduvad järsult ja põhjustavad lehtede kollast värvi.
Porgandijuurtes tekivad kromoplastid leukoplastidest, algul tärkliserikkad, plastiidi stroomas aga kogunevad karotenoidid, mis hiljem kristalliseeruvad. Tärklis kaob, kui karoteeni kontsentratsioon suureneb, plastiidi mass väheneb ja muutub raskesti tuvastatavaks. Kristalliseerunud pigment moodustab suurema osa kromoplastist, nii et kromoplasti kuju määrab lõpuks kristalliseeriva pigmendi kuju ja see on tavaliselt ebakorrapärane: hambuline, poolkuukujuline, nõelakujuline või lamelljas. Plaadid võivad olla kolmnurga, rombi, rööpküliku jne kujuga.

Joonisel on üks vaarika viljalihaga arbuusi rakk vaadatuna läbi valgusmikroskoobi. Rakus on nähtav tsütoplasma, mis koosneb erinevatesse suundadesse venitatud õhukestest filamentidest. Tsütoplasma massiivsemates ahelates paiknevad kromoplasti pigmendi nõelakujulised kristallid. Suurimat kristallide kogunemist täheldatakse südamiku lähedal. Teises karmiinvärvi viljalihaga arbuusi sordis kristalliseerub kromoplasti pigment mitte ainult nõelakujuliste kristallide kujul, vaid ka erineva suurusega lühikeste prismade kujul.
Kromoplastide tähtsust ainevahetuses on väga vähe selgitatud. Sarnaselt leukoplastidele puudub neil fotosünteesivõime, kuna need ei sisalda klorofülli. Kromoplastide kaudne tähtsus seisneb selles, et need määravad lillede ja viljade erksa värvuse, mis meelitab putukaid risttolmlemiseks ja teisi loomi viljade levitamiseks.
Kui leiate vea, tõstke esile mõni tekstiosa ja klõpsake Ctrl+Enter.
Plastiidid on taimerakkude ja mõnede fotosünteetiliste algloomade organellid. Loomadel ja seentel plastiidid puuduvad.
Plastiidid jagunevad mitut tüüpi. Kõige olulisem ja tuntuim on kloroplast, mis sisaldab rohelist pigmenti klorofülli, mis tagab fotosünteesi protsessi.
Muud tüüpi plastiidid on mitmevärvilised kromoplastid ja värvitud leukoplastid. Eraldatud on ka amüloplastid, lipidoplastid, proteinoplastid, mida sageli peetakse leukoplastide sortideks.
Plastiidide tüübid: kloroplastid, kromoplastid, leukoplastid
Kõik plastiidide tüübid on omavahel seotud ühise päritolu või võimaliku vastastikuse muundamise kaudu. Plastiidid arenevad proplastiididest – meristeemiliste rakkude väiksematest organellidest.

Plastiidide struktuur
Enamik plastiide on kahemembraanilised organellid, neil on välimine ja sisemine membraan. Siiski on organisme, mille plastiididel on neli membraani, mis on seotud nende päritolu iseärasustega.
Paljudes plastiidides, eriti kloroplastides, on sisemembraani süsteem hästi arenenud, moodustades selliseid struktuure nagu tülakoidid, grana (tülakoidide virnad), lamellid – piklikud tülakoidid, mis ühendavad naabergrana. Plastiidide sisemist sisu nimetatakse tavaliselt stroomiks. Muuhulgas sisaldab see tärkliseterasid.
Arvatakse, et evolutsiooni käigus ilmusid plastiidid sarnaselt mitokondritele – peremeesrakku viides teise prokarüootse raku, mis on antud juhul võimeline fotosünteesiks. Seetõttu peetakse plastiide poolautonoomseteks organellideks. Nad võivad jaguneda raku jagunemisest sõltumatult, neil on oma DNA, RNA, prokarüootset tüüpi ribosoomid ehk oma valke sünteesiv aparaat. See ei tähenda, et tsütoplasmast pärit valgud ja RNA plastiididesse ei satuks. Osa nende toimimist kontrollivatest geenidest asub just tuumas.
Plastiidide funktsioonid
Plastiidide funktsioonid sõltuvad nende tüübist. Kloroplastid täidavad fotosünteesi funktsiooni. Leukoplastidesse kogunevad varutoitained: tärklis amüloplastides, rasvad elaioplastides (lipidoplastides), valgud proteinoplastides.
Kromoplastid värvivad neis sisalduvate karotenoidpigmentide tõttu erinevaid taimeosi – lilli, vilju, juuri, sügislehti jne. Hele värv on sageli omamoodi signaaliks tolmeldavatele loomadele ning viljade ja seemnete levitajatele.
Taimede degenereeruvates rohelistes osades muutuvad kloroplastid kromoplastideks. Klorofüllipigment hävib, mistõttu ülejäänud pigmendid, hoolimata väikesest kogusest, muutuvad plastiidides märgatavaks ja värvivad sama lehestiku kollakaspunaste toonidega.
Elu arengu koidikul Maal esindasid kõiki rakuvorme bakterid. Nad imesid läbi keha pinna ürgookeanis lahustunud orgaanilist ainet.
Aja jooksul kohanesid mõned bakterid tootma anorgaanilistest orgaanilisi aineid. Selleks kasutasid nad päikesevalguse energiat. Tekkis esimene ökoloogiline süsteem, milles need organismid olid tootjad. Selle tulemusena ilmus Maa atmosfääri nende organismide poolt eraldunud hapnik. Selle abil saate samast toidust palju rohkem energiat ja kasutada lisaenergiat keha ehituse keeruliseks muutmiseks: keha jagamiseks osadeks.
Elu üks olulisi saavutusi on tuuma ja tsütoplasma eraldamine. Tuum sisaldab pärilikku teavet. Südamikku ümbritsev spetsiaalne membraan võimaldas kaitsta juhuslike kahjustuste eest. Vajadusel saab tsütoplasma tuumalt käsklusi, mis suunavad raku elutegevust ja arengut.
Organismid, mille tuum on tsütoplasmast eraldatud, moodustasid tuuma superkuningriigi (nende hulka kuuluvad taimed, seened, loomad).
Nii tekkis ja arenes rakk – taimede ja loomade organiseerimise alus – bioloogilise evolutsiooni käigus.
Isegi palja silmaga ja veelgi parem luubi all on näha, et küpse arbuusi viljaliha koosneb väga väikestest teradest ehk teradest. Need on rakud - väikseimad "tellised", mis moodustavad kõigi elusorganismide, sealhulgas taimede kehad.
Taime elu kulgeb tema rakkude ühine tegevus, luues ühtse terviku. Taimeosade hulkraksusega toimub nende funktsioonide füsioloogiline diferentseerumine, erinevate rakkude spetsialiseerumine sõltuvalt nende asukohast taimekehas.
Taimerakk erineb loomarakust selle poolest, et sellel on tihe kest, mis katab igast küljest sisemise sisu. Lahter ei ole tasane (nagu seda tavaliselt kujutatakse), tõenäoliselt näeb see välja nagu väga väike viaal, mis on täidetud lima sisuga.
Taimeraku ehitus ja funktsioonid
Vaatleme rakku kui organismi struktuurset ja funktsionaalset üksust. Väljaspool on rakk kaetud tiheda rakuseinaga, milles on õhemad lõigud – poorid. Selle all on väga õhuke kile – membraan, mis katab raku sisu – tsütoplasma. Tsütoplasmas on õõnsused - rakumahlaga täidetud vakuoolid. Raku keskel või rakuseina lähedal on tihe keha – tuum koos tuumaga. Tuum on tsütoplasmast eraldatud tuumaümbrisega. Väikesed kehad, plastiidid, on jaotunud kogu tsütoplasmas.
Taimeraku struktuur
Taimeraku organellide ehitus ja funktsioonid
| Organoid | Pilt | Kirjeldus | Funktsioon | Iseärasused |
Rakusein või plasmamembraan | Värvitu, läbipaistev ja väga vastupidav | Läbib rakku ja vabastab rakust aineid. | Rakumembraan on poolläbilaskev |
|
Tsütoplasma | Paks viskoosne aine | See sisaldab kõiki teisi raku osi. | On pidevas liikumises |
|
Tuum (raku oluline osa) | ümmargune või ovaalne | Tagab pärilike omaduste ülekandumise tütarrakkudele jagunemise käigus | Raku keskosa |
|
Sfääriline või ebakorrapärane kuju | Osaleb valkude sünteesis | |||
 | Tsütoplasmast membraaniga eraldatud reservuaar. Sisaldab rakumahla | Varu toitained ja jääkained, mis on rakule ebavajalikud, kogunevad. | Raku kasvades ühinevad väikesed vakuoolid üheks suureks (keskseks) vakuooliks |
|
plastiidid | Kloroplastid | Kasutage päikese valgusenergiat ja looge anorgaanilisest orgaanilisest | Tsütoplasmast topeltmembraaniga eraldatud ketaste kuju |
|
Kromoplastid | Moodustub karotenoidide kuhjumise tulemusena | Kollane, oranž või pruun |
||
 | Leukoplastid | Värvusetud plastiidid | ||
tuumaümbris | Koosneb kahest pooridega membraanist (välimine ja sisemine). | Eraldab tuuma tsütoplasmast | Võimaldab vahetust tuuma ja tsütoplasma vahel |
Raku elusosa on membraaniga piiratud, korrastatud, struktureeritud biopolümeeride ja sisemiste membraanistruktuuride süsteem, mis osaleb metaboolsete ja energiaprotsesside kogumikus, mis säilitavad ja taastoodavad kogu süsteemi tervikuna.
Oluline omadus on see, et rakus ei ole avatud membraane, millel on vabad otsad. Rakumembraanid piiravad alati õõnsusi või piirkondi, sulgedes need igast küljest.

Kaasaegne taimeraku üldistatud diagramm
plasmalemma(välimine rakumembraan) - 7,5 nm paksune ultramikroskoopiline kile, mis koosneb valkudest, fosfolipiididest ja veest. See on väga elastne kile, mis on veega hästi niisutatud ja taastab kiiresti pärast kahjustusi terviklikkuse. Sellel on universaalne struktuur, st tüüpiline kõikidele bioloogilistele membraanidele. Taimerakkudel väljaspool rakumembraani on tugev rakusein, mis loob välise toe ja hoiab raku kuju. See koosneb kiudainetest (tselluloosist), vees lahustumatust polüsahhariidist.
Plasmodesmaat taimerakust on submikroskoopilised tuubulid, mis tungivad läbi membraanide ja on vooderdatud plasmamembraaniga, mis seega katkeb ühest rakust teise. Nende abiga toimub orgaanilisi toitaineid sisaldavate lahuste rakkudevaheline ringlus. Nad edastavad ka biopotentsiaale ja muud teavet.
Poromy nimetatakse aukudeks sekundaarses membraanis, kus rakke eraldavad ainult esmane membraan ja keskmine plaat. Primaarse membraani ja keskmise plaadi piirkondi, mis eraldavad külgnevate rakkude külgnevaid poore, nimetatakse pooride membraaniks või poori sulgevaks kileks. Poori sulgev kile on läbistatud plasmodesmenaalsete tuubulitega, kuid läbivat auku tavaliselt pooridesse ei teki. Poorid hõlbustavad vee ja lahustunud ainete transporti rakust rakku. Naaberrakkude seintes moodustuvad poorid reeglina üksteise vastu.
Raku sein sellel on selgelt piiritletud, suhteliselt paks polüsahhariidne kest. Taime rakusein on tsütoplasma saadus. Selle moodustamisel osalevad aktiivselt Golgi aparaat ja endoplasmaatiline retikulum.
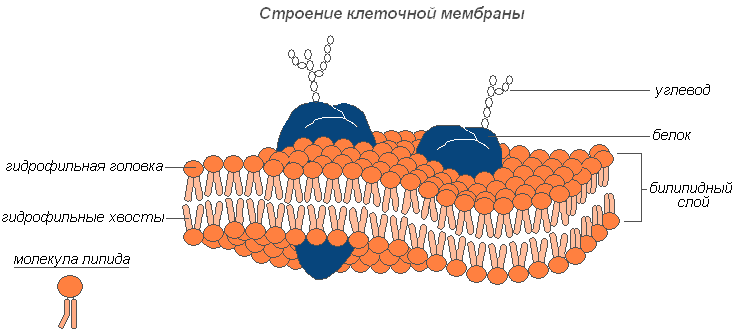
Rakumembraani struktuur
Tsütoplasma aluseks on selle maatriks ehk hüaloplasma, kompleksne värvitu, optiliselt läbipaistev kolloidsüsteem, mis on võimeline pöörduvalt üleminekuks soolilt geelile. Hüaloplasma kõige olulisem roll on ühendada kõik rakustruktuurid ühtseks süsteemiks ja tagada nendevaheline interaktsioon raku ainevahetuse protsessides.
Hüaloplasma(või tsütoplasma maatriks) moodustab raku sisekeskkonna. Koosneb veest ja erinevatest biopolümeeridest (valgud, nukleiinhapped, polüsahhariidid, lipiidid), millest põhiosa moodustavad erineva keemilise ja funktsionaalse spetsiifilisusega valgud. Hüaloplasma sisaldab ka aminohappeid, monosuhkruid, nukleotiide ja muid madala molekulmassiga aineid.
Biopolümeerid moodustavad veega kolloidse keskkonna, mis olenevalt tingimustest võib olla tihe (geeli kujul) või vedelam (sooli kujul) nii kogu tsütoplasmas kui ka selle üksikutes osades. Hüaloplasmas paiknevad mitmesugused organellid ja inklusioonid, mis interakteeruvad üksteisega ja hüaloplasma keskkonnaga. Pealegi on nende asukoht enamasti spetsiifiline teatud rakutüüpidele. Läbi bilipiidmembraani suhtleb hüaloplasma rakuvälise keskkonnaga. Järelikult on hüaloplasma dünaamiline keskkond ja mängib olulist rolli üksikute organellide toimimises ja rakkude kui terviku elutegevuses.
Tsütoplasmaatilised moodustised - organellid
Organellid (organellid) on tsütoplasma struktuurikomponendid. Neil on teatud kuju ja suurus, need on raku kohustuslikud tsütoplasmaatilised struktuurid. Nende puudumisel või kahjustumisel kaotab rakk tavaliselt võime edasi eksisteerida. Paljud organellid on võimelised jagunema ja ise paljunema. Need on nii väikesed, et neid saab näha ainult elektronmikroskoobiga.
Tuum
Tuum on raku kõige nähtavam ja tavaliselt suurim organell. Seda uuris esmakordselt üksikasjalikult Robert Brown 1831. aastal. Tuum tagab raku kõige olulisemad metaboolsed ja geneetilised funktsioonid. See on üsna muutuva kujuga: see võib olla sfääriline, ovaalne, labakujuline, läätsekujuline.
Tuum mängib raku elus olulist rolli. Rakk, millelt tuum on eemaldatud, ei erita enam kesta, lakkab kasvamast ja aineid sünteesimast. Selles intensiivistuvad lagunemis- ja hävimisproduktid, mille tagajärjel see kiiresti sureb. Tsütoplasmast uue tuuma moodustumist ei toimu. Uued tuumad tekivad ainult vana lõhustumise või purustamise teel.
Tuuma sisemine sisu on karüolümf (tuumamahl), mis täidab ruumi tuuma struktuuride vahel. See sisaldab ühte või mitut nukleooli, samuti märkimisväärset hulka DNA molekule, mis on seotud spetsiifiliste valkude - histoonidega.

Tuuma ehitus
nucleolus
Tuum sisaldab sarnaselt tsütoplasmaga peamiselt RNA-d ja spetsiifilisi valke. Selle kõige olulisem ülesanne on see, et selles tekivad ribosoomid, mis teostavad rakus valkude sünteesi.
golgi aparaat
Golgi aparaat on organoid, millel on universaalne jaotus igat tüüpi eukarüootsetes rakkudes. See on mitmetasandiline lamedate membraanikottide süsteem, mis paksenevad piki perifeeriat ja moodustavad vesikulaarseid protsesse. Kõige sagedamini asub see tuuma lähedal.

golgi aparaat
Golgi aparaat sisaldab tingimata väikeste vesiikulite (vesiikulite) süsteemi, mis on kinnitatud paksendatud tsisternidest (ketastest) ja asuvad piki selle struktuuri perifeeriat. Need vesiikulid mängivad spetsiifiliste sektoraalsete graanulite rakusisese transpordisüsteemi rolli ja võivad olla rakuliste lüsosoomide allikad.
Golgi aparaadi funktsioonid seisnevad ka rakusiseste sünteesiproduktide, lagunemissaaduste ja toksiliste ainete rakust väljapoole kogunemises, eraldamises ja vabastamises mullide abil. Raku sünteetilise aktiivsuse saadused, aga ka mitmesugused ained, mis sisenevad rakku endoplasmaatilise retikulumi kanalite kaudu keskkonnast, transporditakse Golgi aparaati, kogunevad sellesse organoidi ja sisenevad seejärel kujul tsütoplasmasse. tilkadest või teradest ja neid kasutab rakk ise või eritub. Taimerakkudes sisaldab Golgi aparaat ensüüme polüsahhariidide sünteesiks ja polüsahhariidmaterjali ennast, mida kasutatakse rakuseina ehitamiseks. Arvatakse, et see on seotud vakuoolide moodustumisega. Golgi aparaat sai nime Itaalia teadlase Camillo Golgi järgi, kes avastas selle esmakordselt 1897. aastal.
Lüsosoomid
Lüsosoomid on väikesed vesiikulid, mis on piiratud membraaniga, mille põhiülesanne on rakusisene seedimine. Lüsosomaalse aparaadi kasutamine toimub taime seemne idanemise ajal (varu toitainete hüdrolüüs).
Lüsosoomi struktuur
mikrotuubulid
Mikrotuubulid on membraansed supramolekulaarsed struktuurid, mis koosnevad spiraalselt või sirgete ridadena paigutatud valgugloobulitest. Mikrotuubulid täidavad valdavalt mehaanilist (motoorset) funktsiooni, tagades rakuorganellide liikuvuse ja kontraktiilsuse. Asudes tsütoplasmas, annavad nad rakule kindla kuju ja tagavad organellide ruumilise paigutuse stabiilsuse. Mikrotuubulid hõlbustavad organellide liikumist asukohtadesse, mis on määratud raku füsioloogiliste vajadustega. Märkimisväärne osa neist struktuuridest paikneb plasmalemmas, rakumembraani lähedal, kus nad osalevad taimerakumembraanide tselluloosi mikrofibrillide moodustamises ja orientatsioonis.

Mikrotuubulite struktuur
Vacuool
Vakuool on taimerakkude kõige olulisem komponent. See on omamoodi õõnsus (reservuaar) tsütoplasma massis, mis on täidetud mineraalsoolade, aminohapete, orgaaniliste hapete, pigmentide, süsivesikute vesilahusega ja eraldatud tsütoplasmast vaakummembraaniga - tonoplastiga.
Tsütoplasma täidab kogu sisemise õõnsuse ainult kõige nooremates taimerakkudes. Raku kasvuga muutub oluliselt tsütoplasma algselt pideva massi ruumiline paigutus: sellesse tekivad väikesed rakumahlaga täidetud vakuoolid ja kogu mass muutub käsnjaks. Rakkude edasisel kasvul üksikud vakuoolid ühinevad, surudes tsütoplasmaatilised kihid perifeeriasse, mille tulemusena on tekkinud rakus tavaliselt üks suur vakuool ning tsütoplasma koos kõigi organellidega paikneb membraani läheduses.
Veeslahustuvad orgaanilised ja mineraalsed vakuoolide ühendid määravad elusrakkude vastavad osmootsed omadused. See teatud kontsentratsiooniga lahus on omamoodi osmootne pump rakku kontrollitud tungimiseks ja vee, ioonide ja metaboliitide molekulide vabastamiseks sellest.
Koos tsütoplasma kihi ja selle membraanidega, mida iseloomustavad poolläbilaskvusomadused, moodustab vakuool tõhusa osmootse süsteemi. Osmootselt määratud on sellised elusate taimerakkude näitajad nagu osmootne potentsiaal, imemisjõud ja turgorirõhk.

Vakuooli struktuur
plastiidid
Plastiidid on suurimad (tuuma järel) tsütoplasmaatilised organellid, mis on omased ainult taimerakkudele. Neid ei leidu ainult seentes. Plastiidid mängivad olulist rolli ainevahetuses. Need on tsütoplasmast eraldatud topeltmembraanimembraaniga ning mõnel nende tüübil on hästi arenenud ja korrastatud sisemembraanide süsteem. Kõik plastiidid on sama päritoluga.
Kloroplastid- fotoautotroofsete organismide levinumad ja funktsionaalselt olulisemad plastiidid, mis viivad läbi fotosünteesiprotsesse, mis lõppkokkuvõttes viivad orgaaniliste ainete moodustumiseni ja vaba hapniku vabanemiseni. Kõrgemate taimede kloroplastidel on keeruline sisemine struktuur.

Kloroplasti struktuur
Kloroplastide suurused erinevates taimedes ei ole samad, kuid keskmiselt on nende läbimõõt 4-6 mikronit. Kloroplastid on võimelised liikuma tsütoplasma liikumise mõjul. Lisaks täheldatakse valgustuse mõjul amööboid-tüüpi kloroplastide aktiivset liikumist valgusallika poole.
Klorofüll on kloroplastide peamine aine. Tänu klorofüllile on rohelised taimed võimelised kasutama valgusenergiat.
Leukoplastid(värvitud plastiidid) on selgelt märgistatud tsütoplasma kehad. Nende mõõtmed on mõnevõrra väiksemad kui kloroplastide suurused. Ühtlasemad ja nende kuju, lähenedes kerakujulisele.

Leukoplasti struktuur
Neid leidub epidermise, mugulate, risoomide rakkudes. Valgustatuna muutuvad need väga kiiresti kloroplastideks koos vastava sisestruktuuri muutumisega. Leukoplastid sisaldavad ensüüme, mille abil sünteesitakse fotosünteesi käigus tekkivast liigsest glükoosist tärklis, millest suurem osa ladestub tärkliseteradena säilituskudedesse või -organitesse (mugulad, risoomid, seemned). Mõnes taimes ladestuvad rasvad leukoplastidesse. Leukoplastide reservfunktsioon avaldub aeg-ajalt säilitusvalkude moodustumisel kristallide või amorfsete inklusioonide kujul.
Kromoplastid enamasti on need kloroplastide, aeg-ajalt leukoplastide derivaadid.

Kromoplasti struktuur
Kibuvitsamarjade, paprikate, tomatite valmimisega kaasneb viljaliharakkude kloro- või leukoplastide muundumine karotenoidideks. Viimased sisaldavad valdavalt kollaseid plastiidpigmente – karotenoide, mis laagerdumisel neis intensiivselt sünteesitakse, moodustades värvilisi lipiiditilku, tahkeid kerakesi või kristalle. Klorofüll hävib.
Mitokondrid
Mitokondrid on organellid, mida leidub enamikus taimerakkudes. Neil on muutuva kujuga pulgad, terad, niidid. Need avastas 1894. aastal R. Altman valgusmikroskoobi abil ning hiljem uuriti sisestruktuuri kasutades elektroonilist.

Mitokondrite struktuur
Mitokondritel on kahe membraaniga struktuur. Välimine membraan on sile, sisemine moodustab erineva kujuga väljakasvu - tuubuleid taimerakkudes. Mitokondrite sees olev ruum on täidetud poolvedela sisaldusega (maatriksiga), mis sisaldab ensüüme, valke, lipiide, kaltsiumi- ja magneesiumisooli, vitamiine, aga ka RNA-d, DNA-d ja ribosoome. Mitokondrite ensümaatiline kompleks kiirendab keeruka ja omavahel seotud biokeemiliste reaktsioonide mehhanismi tööd, mille tulemusena moodustub ATP. Nendes organellides on rakud varustatud energiaga - toitainete keemiliste sidemete energia muundatakse rakulise hingamise käigus ATP kõrge energiasisaldusega sidemeteks. Just mitokondrites toimub süsivesikute, rasvhapete, aminohapete ensümaatiline lagunemine koos energia vabanemisega ja selle järgneva muundamisega ATP energiaks. Kogunenud energia kulub kasvuprotsessidele, uutele sünteesidele jne. Mitokondrid paljunevad jagunemise teel ja elavad umbes 10 päeva, misjärel nad hävivad.
Endoplasmaatiline retikulum
Endoplasmaatiline retikulum - tsütoplasmas paiknev kanalite, tuubulite, vesiikulite, tsisternide võrgustik. See on 1945. aastal inglise teadlase K. Porteri poolt avatud ultramikroskoopilise struktuuriga membraanide süsteem.

Endoplasmaatilise retikulumi struktuur
Kogu võrk on integreeritud tuumaümbrise välimise rakumembraaniga üheks tervikuks. Eristage ER siledat ja karedat, ribosoome kandvat. Sileda EPS-i membraanidel on ensüümsüsteemid, mis osalevad rasvade ja süsivesikute ainevahetuses. Seda tüüpi membraan valitseb reservainete (valgud, süsivesikud, õlid) rikastes seemnerakkudes, granuleeritud ER-i membraanile kinnituvad ribosoomid ja valgumolekuli sünteesi käigus sukeldub ER-i polüpeptiidahel koos ribosoomidega. kanal. Endoplasmaatilise retikulumi funktsioonid on väga mitmekesised: ainete transport nii rakusiseselt kui ka naaberrakkude vahel; raku jagunemine eraldi sektsioonideks, milles toimuvad samaaegselt erinevad füsioloogilised protsessid ja keemilised reaktsioonid.
Ribosoomid
Ribosoomid on mittemembraansed rakulised organellid. Iga ribosoom koosneb kahest ebavõrdse suurusega osakesest ja selle saab jagada kaheks fragmendiks, mis säilitavad pärast tervikuks ribosoomiks ühinemist võimet valku sünteesida.

Ribosoomi struktuur
Ribosoomid sünteesitakse tuumas, seejärel lahkuvad sellest, sisenedes tsütoplasmasse, kus need kinnituvad endoplasmaatilise retikulumi membraanide välispinnale või paiknevad vabalt. Sõltuvalt sünteesitava valgu tüübist võivad ribosoomid toimida üksi või ühineda kompleksideks – polüribosoomideks.
Loe ka...
- Kas ülikooli astumisel on soodustusi paljulapselistele peredele Paljulapseliste perede toetuste nimekiri
- Verejooksu põhjused raseduse alguses: mida teha?
- Mida teha, kui lapsel on kõndimisel valu reie piirkonnas: põhjused, ravi ja rahvapärased abinõud
- Mida teha ja kuidas ravida vesist kõhulahtisust lapsel?


















